miR-155
[4][5][6][7][8][9] Exogenous molecular control in vivo of miR-155 expression may inhibit malignant growth,[10][11] viral infections,[12] and enhance the progression of cardiovascular diseases.
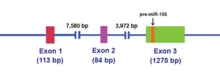
[14] The MIR155HG RNA transcript does not contain a long open reading frame (ORF), however, it does include an imperfectly base-paired stem loop that is conserved across species.
[15] Once miR-155 pri-miRNA is transcribed, this transcript is cleaved by the nuclear microprocessor complex, of which the core components are the RNase III type endonuclease Drosha and the DiGeorge critical region 8 (DGCR8) protein,[16][17] to produce a 65 nucleotide stem-loop precursor miRNA (pre-mir-155) (see Figure 2).
[2] Northern blot analysis found that miR-155 pri-miRNA was abundantly expressed in the human spleen and thymus and detectable in the liver, lung, and kidney.
A comprehensive list of miR-155-5p/mRNA targets that were experimentally authenticated by both the demonstration of endogenous transcript regulation by miR-155-5p and validation of the miR-155-5p seed sequence through a reporter assay was recently assembled.
[31] Additionally, Hu et al.[32] demonstrated that the homeobox protein, HOXA9, regulated MIR155HG expression in myeloid cells and that this miRNA played a functional role in hematopoiesis.
These investigators found that forced expression of miR-155-5p in bone marrow cells resulted in a ~50% decrease in SPI1 (i.e. PU.1),[32] a transcription factor and a regulator of myelopoiesis,[33] and a validated target of this miRNA.
[34] Additionally, mice deficient in pre-mir-155 showed clear defects in lymphocyte development and generation of B- and T-cell responses in vivo.
[37][38] Taken together, these results strongly suggest that miR-155-5p is an essential molecule in the control of several aspects of hematopoiesis including myelopoiesis, erythropoiesis, and lymphopoiesis.
The innate immune system constitutes the first line of defense against invading pathogens and is regarded as the major initiator of inflammatory responses.
[47] In the context of viral infection vesicular stomatitis virus (VSV) challenge of murine peritoneal macrophages was reported to result in miR-155-5p over-expression via a retinoic acid-inducible gene I/JNK/NF-κB–dependent pathway.
[48] Support for a role of AP-1 in MIR155HG activation comes from studies using stimuli relevant to viral infection such as TLR3 ligand poly(I:C) or interferon beta (IFN-β).
These include inositol polyphosphate-5-phosphatase (INPP5D also denoted SHIP1) and suppressor of cytokine signaling 1 (SOCS1), suppression of which promotes cell survival, growth, migration, and anti-pathogen responses.
[48][53][54][55] Besides supporting the activation of defense pathways miR-155-5p may also limit the strength of the resulting NF-κB dependent inflammatory response,[52] suggesting varying functions of miR-155 at different stages of inflammation.
There is evidence that miR-155 participates in cascades associated with cardiovascular diseases and hypertension, and was also found to be implicated in immunity, genomic instability, cell differentiation, inflammation, virus associated infections, cancer, and diabetes mellitus.
[56] Protective roles of miR-155 may arise in response to its action on silencing genes thereby regulating their expression time, mutations in miR-155 target site deny it the optimal access necessary to bring about gene silencing, leading to over abundance of delinquent activities that may go malignant, for example, miR-155 role as a protective agent against predisposition to B Cell associated malignancies is emphasized by maintaining the balance of Activation-Induced Cytidine Deaminase (AID) enzyme.
[4] miR-155 is involved in immunity by playing key roles in modulating humoral and innate cell-mediated immune responses, for example, In miR-155 deficient mice, immunological-memory is impaired; making it fall prey to repetitive bouts of invasions by the same pathogen (Rodriguez et al. 2007), maturation and specificity of miR-155-deficient B-lymphocytes are impaired since the process relies on AID enzyme which has a miR-155 target in its 3′ UTR end.
The change in IgG1 levels maybe explained by the fact that it is a target for miR-155 in B cells, the protein-encoding mRNA for the transcriptional regulator Pu.1-protein, elevation of Pu.1 protein predisposes defective IgG1 production.
[4] In multiple sclerosis, increased expression of mir-155 has also been measured in peripheral and CNS-resident myeloid cells, including circulating blood monocytes and activated microglia.