Pain in fish
[2][3][4] In 1789, the British philosopher and social reformist, Jeremy Bentham, addressed in his book An Introduction to the Principles of Morals and Legislation the issue of our treatment of animals with the following often quoted words: "The question is not, Can they reason?
[8] In his interactions with scientists and other veterinarians, Rollin was regularly asked to "prove" that animals are conscious, and to provide "scientifically acceptable" grounds for claiming that they feel pain.
Convergent evidence indicates that non-human animals have the neuroanatomical, neurochemical, and neurophysiological substrates of conscious states along with the capacity to exhibit intentional behaviors.
She stated that fish demonstrate pain-related changes in physiology and behaviour, that are reduced by painkillers, and they show higher brain activity when painfully stimulated.
[22] At the same time as the investigations using arthritic rats, studies were published showing that birds with gait abnormalities self-select for a diet that contains carprofen, a human analgesic.
[39] This is the ability to detect noxious stimuli which evoke a reflex response that rapidly moves the entire animal, or the affected part of its body, away from the source of the stimulus.
However, nociceptive responses can be so subtle in prey animals that trained (human) observers cannot perceive them, whereas natural predators can and subsequently target injured individuals.
There have been several published lists of criteria for establishing whether non-human animals experience pain, e.g.[41][42] Some criteria that may indicate the potential of another species, including fishes, to feel pain include:[42] The adaptive value of nociception is obvious; an organism detecting a noxious stimulus immediately withdraws the limb, appendage or entire body from the noxious stimulus and thereby avoids further (potential) injury.
[47][48] The study was designed to determine whether nociceptors were present in the trigeminal nerve on the head of the trout and to observe the physiological and behavioural consequences of prolonged noxious stimulation.
[52] Rainbow trout (Oncorhynchus mykiss) have polymodal nociceptors on the face and snout that respond to mechanical pressure, temperatures in the noxious range (> 40 °C), and 1% acetic acid (a chemical irritant).
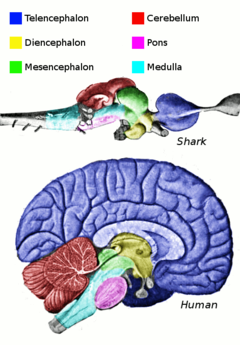
[64] The central nervous system (CNS) of fish contains a spinal cord, medulla oblongata, and the brain, divided into telencephalon, diencephalon, mesencephalon and cerebellum.
[67] Microarray analysis of gene expression shows the brain is active at the molecular level in the forebrain, midbrain and hindbrain of common carp and rainbow trout.
[77] All four of the main opioid receptor types (delta, kappa, mu, and NOP) are conserved in vertebrates, even in primitive jawless fishes (agnathastoma).
[78][79] Pre-treatment with morphine (an analgesic in humans and other mammals) has a dose-dependent anti-nociceptive effect[80] and mitigates the behavioural and ventilation rate responses of rainbow trout to noxious stimuli.
[71] Injection of acetic acid into the lips of rainbow trout causes a reduction in their natural neophobia (fear of novelty); this is reversed by the administration of morphine.
In a study on the efficacy of three types of analgesic, buprenorphine (an opioid), carprofen (a non-steroidal anti-inflammatory drug) and lidocaine (a local anaesthetic), ventilation rate and time to resume feeding were used as pain indicators.
Buprenorphine had limited impact on the fish's response, carprofen ameliorated the effects of noxious stimulation on time to resume feeding, however, lidocaine reduced all the behavioural indicators.
[92] Nile tilapia (Oreochromis niloticus), in response to a tail fin clip, increase their swimming activity and spend more time in the light area of their tank.
[93] Since this initial work, Sneddon and her co-workers have shown that rainbow trout, common carp and zebrafish experiencing a noxious stimulation exhibit rapid changes in physiology and behavior that persist for up to 6 hours and thus are not simple reflexes.
The researchers suggested the findings of the motivational changes and trade-offs provide evidence for central processing of pain rather than merely showing a nociceptive reflex.
Examples include the ability to recognise social companions, avoidance (for some months or years) of places where they encountered a predator or were caught on a hook and forming mental maps.
[102] Scientists have also proposed that in conjunction with argument-by-analogy, criteria of physiology or behavioural responses can be used to assess the possibility of non-human animals perceiving pain.
[104][105][106] Other societal implications of fish experiencing pain include acute and chronic exposure to pollutants, commercial and sporting fisheries (e.g. injury during trawling, tagging/fin clipping during stock assessment, tissue damage, physical exhaustion and severe oxygen deficit during capture, pain and stress during slaughter, use of live bait), aquaculture (e.g. tagging/fin clipping, high stocking densities resulting in increased aggression, food deprivation for disease treatment or before harvest, removal from water for routine husbandry, pain during slaughter), ornamental fish (e.g. capture by sub-lethal poisoning, permanent adverse physical states due to selective breeding), scientific research (e.g. genetic-modification) may have detrimental effects on welfare, deliberately-imposed adverse physical, physiological and behavioural states, electrofishing, tagging, fin clipping or otherwise marking fish, handling procedures which may cause injury.
[46][107] Browman et al.[108] suggest that if the regulatory environment continues on its current trajectory (adding more aquatic animal taxa to those already regulated), activity in some sectors could be severely restricted, even banned.
He points out that the alternative explanation is that elasmobranchs have lost the ability of nociception, and one would have to come up with a very convincing argument for the adaptive value of such a loss in a single taxon in the entire animal kingdom.
[117] Professor Broom of Cambridge University, submits that feeling pain gives active complex vertebrates a selective advantage through learning and responding, allowing them to survive in their environment.
There is also evidence from activation studies showing brainstem mediated feelings in normal humans and foetal withdrawal responses to noxious stimulation but prior to development of the cortex.
[131] In papers published in 2017 and 2018, Michael Woodruff[132][133] summarized a significant number of research articles that, in contradiction to the conclusions of Rose and Key, strongly support the hypothesis that the neuroanatomical organization of the fish pallium and its connections with subpallial structures, especially those with the preglomerular nucleus and the tectum, are complex enough to be analogous to the circuitry of the cortex and thalamus assumed to underlie sentience in mammals.
He added neurophysiological and behavioral data to these anatomical observations that also support the hypothesis that the pallium is an important part of the hierarchical network proposed by Feinberg and Mallatt to underlie consciousness in fishes.
[60] In 2018, Sneddon, Donald Broom, Culum Brown and others, published a paper that found that despite the empirical proof, sceptics still deny anything beyond reflex responses in fishes and state that they are incapable of complex cognitive abilities.