Polydnaviriformidae
[2][3] Little or no sequence homology exists between BV and IV, suggesting that the two genera have been evolving independently for a long time.
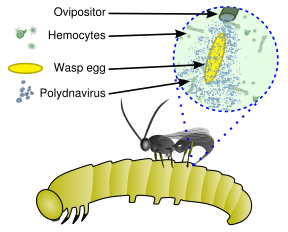
The virus only replicates in a particular part of the ovary, called the calyx, of pupal and adult female wasps.
Many parasitoids that do not use PDVs inject proteins that provide many of the same functions, that is, a suppression of the immune response to the parasite egg.
In this model, the braconid and ichneumonid wasps packaged genes for these functions into the viruses—essentially creating a gene-transfer system that results in the caterpillar producing the immune-suppressing factors.
Following integration, the genes responsible for virus replication and the capsids were (eventually) no longer included in the PDV genome.
[14] IV has a less clear origin: although earlier reports found a protein p44/p53 with structural similarities to ascovirus, the link was not confirmed in later studies.
[15] As a result, the current opinion is that IV originated from a yet-unidentified novel viral family,[13] with a weak link to the NCLDVs.
An encapsulated body can also be melanised in order to asphyxiate it, thanks to another type of hemocyte, which uses the phenoloxidase pathway to produce melanin.
[18] PolyDNAvirus protect the hymenopteran larvae from the host immune system, acting at different levels.
VLPs can be compared to PolyDNAvirus because they are secreted in the same way, and they both act to protect the larvae against the host's immune system.
This link produces the name Venturia canescens endogenous nudivirus (VcENV), an alphanudivirus closely related to NlENV found in Nilaparvata lugens.
[28] MicroRNA are small RNA fragments produced in the host cells thanks to a specific enzymatic mechanism.
This phenomenon is known as PTGS (for post transcriptional gene silencing)[29] or RNAi (RNA interference.)