RIG-I-like receptor
[10][11] LGP2 binds to blunt-ended double-stranded RNA of variable length,[12][13] and also to RNA-bound MDA5 to regulate filament formation.
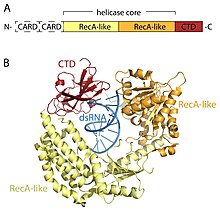
[16] In addition to the helicase core and CTD, RIG-I and MDA5 have two N-terminal CARD (caspase active recruitment domains) that are essential to the initiation of downstream signaling.
[14][17][18][19][20][15][21] In uninfected cells that are absent of viral RNA RIG-I exists in an inactive conformation in which the CARD domains are masked due to their interaction with the CTD.
[23] As a safeguard for RLR activation, the exposed RIG-I and MDA5 CARDs can undergo post-translational modifications (e.g. ubiquitination, phosphorylation) that either positively or negatively regulate downstream signaling.
This binding event is essential to signaling as it causes MAVS to form large functional aggregates in which TRAF3 (TNF receptor-associated factor 3) and subsequently the IKKε/TBK1 (I-kappa-B kinase-epsilon/TANK-binding kinase 1) complex are recruited.
This leads to the induction of hundreds of interferon stimulated genes (ISGs) that amplify the IFN response.
As prolonged IFN production is linked to human disease RLR signaling must be tightly regulated.
Most famously the RIG-I CARD domain is phosphorylated by protein kinase C-α (PKC-α) and PKC-β in the resting state to negatively regulate signaling.
This article was adapted from the following source under a CC BY 4.0 license (2019) (reviewer reports): Natalie Borg (2019).