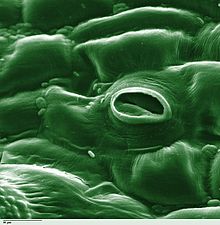
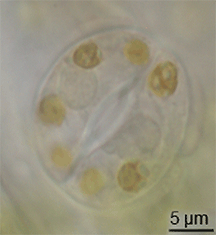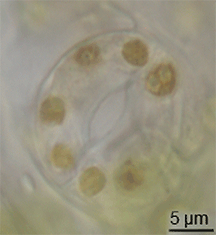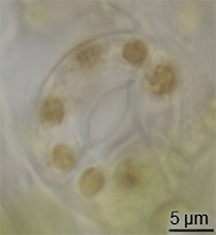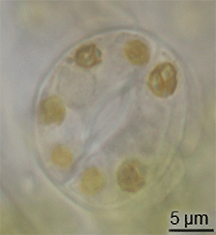
Stoma
: stomates), is a pore found in the epidermis of leaves, stems, and other organs, that controls the rate of gas exchange between the internal air spaces of the leaf and the atmosphere.
[1] Air, containing oxygen, which is used in respiration, and carbon dioxide, which is used in photosynthesis, passes through stomata by gaseous diffusion.
Stomata are present in the sporophyte generation of the vast majority of land plants, with the exception of liverworts, as well as some mosses and hornworts.
[5] Ordinarily, carbon dioxide is fixed to ribulose 1,5-bisphosphate (RuBP) by the enzyme RuBisCO in mesophyll cells exposed directly to the air spaces inside the leaf.
Narrower stomatal apertures can be used in conjunction with an intermediary molecule with a high carbon dioxide affinity, phosphoenolpyruvate carboxylase (PEPcase).
plants (crassulacean acid metabolism, after the family Crassulaceae, which includes the species in which the CAM process was first discovered) open their stomata at night (when water evaporates more slowly from leaves for a given degree of stomatal opening), use PEPcase to fix carbon dioxide and store the products in large vacuoles.
The following day, they close their stomata and release the carbon dioxide fixed the previous night into the presence of RuBisCO.
However, most plants do not have CAM and must therefore open and close their stomata during the daytime, in response to changing conditions, such as light intensity, humidity, and carbon dioxide concentration.
[6] When the roots begin to sense a water shortage in the soil, abscisic acid (ABA) is released.
This allows scientists to investigate how stomata respond to changes in environmental conditions, such as light intensity and concentrations of gases such as water vapor, carbon dioxide, and ozone.
The inverse of r is conductance to water vapor (g), so the equation can be rearranged to[12] and solved for g:[12] Photosynthetic CO2 assimilation (A) can be calculated from where Ca and Ci are the atmospheric and sub-stomatal partial pressures of CO2 respectively[clarification needed].
These scientific instruments measure the amount of water vapour leaving the leaf and the vapor pressure of the ambient air.
[13][14] There is little evidence of the evolution of stomata in the fossil record, but they had appeared in land plants by the middle of the Silurian period.
[16] However, the evolution of stomata must have happened at the same time as the waxy cuticle was evolving – these two traits together constituted a major advantage for early terrestrial plants.
One that is widely used is based on the types that Julien Joseph Vesque introduced in 1889, was further developed by Metcalfe and Chalk,[23] and later complemented by other authors.
[28] However, pathogenic bacteria applied to Arabidopsis plant leaves can release the chemical coronatine, which induce the stomata to reopen.
Guard cell protoplasts swell under blue light provided there is sufficient availability of potassium.
[32] Zeaxanthin in guard cells acts as a blue light photoreceptor which mediates the stomatal opening.
[33] Stomatal density and aperture (length of stomata) varies under a number of environmental factors such as atmospheric CO2 concentration, light intensity, air temperature and photoperiod (daytime duration).
Low humidity stresses guard cells causing turgor loss, termed hydropassive closure.
Hydroactive closure is contrasted as the whole leaf affected by drought stress, believed to be most likely triggered by abscisic acid.
[30] The gene HIC (high carbon dioxide) encodes a negative regulator for the development of stomata in plants.
[40] The existence of a feedback mechanism results a phenotypic plasticity in response to [CO2]atm that may have been an adaptive trait in the evolution of plant respiration and function.