Thalassiosira
[6] The genus was then characterized with transmission electron microscopy in the 1950s and scanning electron microscopy in the 1960s, which led to an enhanced understanding of the defining morphological features of the genus and the subsequent recognition of over 100 species [7] A species of Thalassiosira, T. pseudonana, was selected as the first marine eukaryotic phytoplankton to undergo whole genome sequencing, due to its small 34Mb genome.
Whole genome sequencing, transcriptomics and proteomics of T. pseudonana have revealed novel pathways for silicon biogenesis, phosphorus stress response, intracellular transport and metabolism in marine diatoms.
[5] Thalasiosira harbor several discoid plastids and a circular valve, which contains pores arranged in rows or arcs, opening outwards.
[12] Species in this genus are also capable of assembling defensive threads against zooplankton, allowing them to survive the predation that normally keeps phytoplankton blooms in check.
Iron concentrations, temperature and macronutrient availability have been identified as important factors for the composition of Thalassiosira species communities in marine waters.
[1] Phylogenetic studies based on 18S rRNA genes have revealed several clades within Thalassiosira, an overall paraphyletic group within the family Thalassiosirales.
A surprising finding from the T. pseudonana genome was the presence of genes encoding enzymes for a complete urea cycle, which was unprecedented in a photosynthetic eukaryote.
Curiously, the T. pseudonana urea cycle feeds into other metabolic pathways, which contribute to protein biosynthesis and possibly energy storage.
[16] In sexual reproduction, sperm and egg, which can arise from the same cell, fuse to form a diploid zygote, which is referred to as an auxospore.
Specifically, T. weissflogii has been shown to reduce its silica synthesis while upregulating the production of triacylglycerols when cultured in nitrogen-limiting conditions.
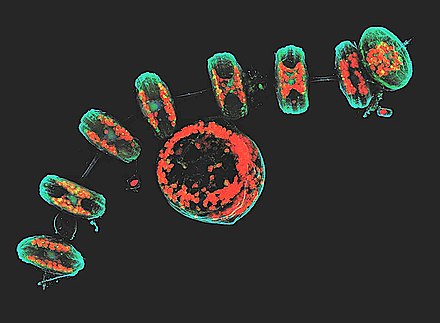
This confocal image shows the diatoms' cell wall (cyan), chloroplasts (red), DNA (blue), membranes and organelles (green).

