Achelousaurus
[2] In the spring of 1985, paleontologist John "Jack" R. Horner was informed that he would no longer be allowed to exploit the Willow Creek site, where he had studied the Maiasaura Egg Mountain nesting colony for six years.
Horner had always been intrigued by the field diaries of Charles Whitney Gilmore who had reported the discovery of dinosaur eggs at Landslide Butte in 1928, but never published on them.
Part of the discoveries made on this occasion was an additional horned dinosaur skull, specimen MOR 492, that later would be referred to (i.e., formally assigned to) Rubeosaurus, the genus name in 2010 given to Styracosaurus ovatus.
[8][9] During the field season of 1987 (early July), volunteer Sidney M. Hostetter located another horned dinosaur skull near the Canyon Bone Bed, specimen MOR 485.
[12] In the summer of 1989, graduate student Scott D. Sampson joined the team, wanting to study the function of the frill display structures in horned dinosaurs.
[13] At the end of June 1989, Horner, his son Jason and his head preparator Carrie Ancell discovered horned dinosaur specimen MOR 591, a subadult skull and partial postcranial skeleton, near the Blacktail Creek.
[14] It was initially assumed that all the horned dinosaur material recovered by the expeditions could be assigned to a single "styracosaur" species distinct from Styracosaurus albertensis, as the fossils represented a limited geological time period, then estimated at half a million years.
This species was informally named "Styracosaurus makeli" in honor of Bob Makela, who had died in a traffic accident just days before the discovery of specimen MOR 485.
He still thought that the fossil material had been part of a single population but concluded that this had developed over time as a chronospecies evolving into a series of subsequent taxa.
In 1992, Horner, David Varricchio, and Mark Goodwin published an article in Nature based on the six-year field study of sediments and dinosaurs from Montana.
This allusion is a reference to the supposedly transitional traits of the dinosaur and the characteristic loss of horns through ontogenetic and phylogenetic development, and thus through individual change and evolution.
[26] The holotype specimen MOR 485 was collected by Hostetter and Ray Rogers[27] from the Landslide Butte Field Area about 40 km (25 mi) northwest of Cut Bank.
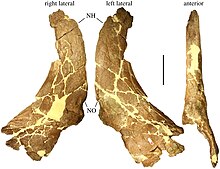
In 1995 Sampson described it as the partial skull of an adult animal including the nasal and supraorbital (region above the eye socket) bosses (roundish protuberances instead of horns), and the parietal bones.
[8] The subadult specimen MOR 591 was assigned to Achelousaurus in 1995 and henceforward, but in 2021, John Wilson and Jack Scannella stated that it could also possibly belong to Einiosaurus.
Achelousaurus exhibited the build of derived ("advanced") centrosaurines, which are typified by short brow horns or bosses, combined with elaborate frill spikes.
During the 1990s, it was increasingly understood that such spikes on the parietals were not random growths but specific traits that could be used to determine the evolution of horned dinosaurs, if only it could be analyzed how they corresponded among species.
[22] In Achelousaurus these epoccipitals, which start as separate skin ossifications or osteoderms, fuse with the underlying frill bone to form spikes,[53] at least in the third position.
[55] In 1992, the study by Horner et al. tried to account for the fact that within a limited geological period of time (about half a million years) there had been a quick succession of animal communities in the upper Two Medicine Formation.
[56] The animals were living on a narrow strip on the east-coast of Laramidia, bordering the Western Interior Seaway and constrained in the west by the 3 to 4 kilometres (2 to 2.5 mi) high proto-Rocky Mountains.
[22] Subsequent studies have sought to determine the precise relationships within this part of the evolutionary tree, with conflicting results regarding the question whether Styracosaurus albertensis or Einiosaurus might have been in the direct line of ascent to Achelousaurus.
[8] In 2011, a subsequent study by Andrew T. McDonald in this respect replicated the outcome of his previous one,[62] as did a publication by Andre Farke et al.[63] In 2017, J.P. Wilson and Ryan further complicated the issue, concluding that MOR 492 ("Taxon A") was not referable to Rubeosaurus and announcing that yet another genus would be named for it.
The bone structure indicates that the bosses were covered by cornified pads as in modern muskoxen, suggesting dominance fights similar to those of members of the Caprinae subfamily.
The bosses would have evolved for actual combat, part of a social selection in which individuals competed for scarce resources such as mates, food and breeding grounds.
[39] Achelousaurus is known from the Two Medicine Formation, which preserves coastal sediments dating from the Campanian stage of the Late Cretaceous Period, between 83 and 74 million years ago.
[78] The brown paleosol in which the horned dinosaurs were found – a mixture of clay and coalified wood fragments – resembles that of modern seasonally dry swamps.
[79] Achelousaurus ate much smaller plants, though: a 2013 study determined that ceratopsid herbivores on Laramidia were restricted to feeding on vegetation with a height of 1 m (3.5 ft) or lower.
As proven by tooth marks, horned dinosaur fossils in the Landslide Butte Field Area had been scavenged by a large theropod predator, which Rogers suggested were Albertosaurus.
Its intermediate anagenetic position suggests that Achelousaurus shared its habitat with forms roughly found in the middle or at the end of the time range of its formation.
In addition, remains of other indeterminate and dubious centrosaurines, including Brachyceratops, are known from the formation and though they may represent younger stages of the three valid genera, this is not possible to demonstrate.
[45][52] Whereas Horner assumed that Einiosaurus and Achelousaurus were separate in time, in 2010 Donald M. Henderson considered it possible that at least their descendants or ancestors were overlapping or sympatric and thus would have competed for food sources unless there had been niche partitioning.