Actin assembly-inducing protein
It has been shown that ActA is not only necessary but also sufficient to induce motility of bacteria in the absence of other bacterial factors.
To determine whether actA itself, plcB or other co-transcribed downstream regions are involved in actin assembly, mutations in the appropriate genes were generated.
[8] Actin filament assembly generates the force that pushes the bacterium in the mammalian host cytoplasm forward.
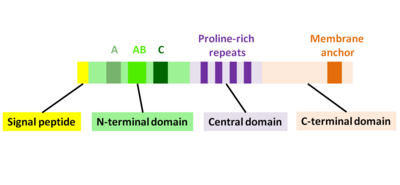
[7] The actin monomer-binding region of ActA has functional properties like the WASP-Homology-2 (WH2) or V domain, but differs in the sequence.
The central proline-rich region of ActA is crucial for ensuring efficient bacterial motility.
These regions mimic those of the host cell cytoskeletal protein zyxin, vinculin and palladin, known to associate with focal adhesions or stress fibers.
[16] The vasodilator-stimulated phosphoprotein (VASP) can bind through its Ena/VASP homology 1 domain (EVH1 domain) to the central proline-rich region and recruits profilin, an actin monomer binding protein, which itself promotes polymerization at barbed ends of actin filaments.
VASP has been shown to reduce the frequency actin-Y-branches in vitro and thus increases the proportion of filaments which are organized in a parallel alignment in comet tails.
[18][19] The C-terminal domain of ActA has a hydrophobic region which anchors the protein in the bacterial membrane.
The Arp2/3 complex is only localized near the bacterial surface and thus it is assumed that a more frequent Arp2/3 complex-independent elongation occurs.