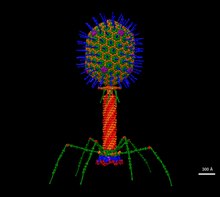
Escherichia virus T4
Dating back to the 1940s and continuing today, T-even phages are considered the best studied model organisms.
[6] When packaged, the concatemer is cut at unspecific positions of the same length, leading to several genomes that represent circular permutations of the original.
[11] The structure of the 6 megadalton T4 baseplate that comprises 127 polypeptide chains of 13 different proteins (gene products 5, 5.4, 6, 7, 8, 9, 10, 11, 12, 25, 27, 48 and 53) has recently been described in atomic detail.
Maintaining an appropriate balance in the amounts of each of these proteins produced during viral infection appears to be critical for normal phage T4 morphogenesis.
This unravels the short tail fibers (STF) that bind irreversibly to the E. coli cell surface.
The remaining part of the membrane is degraded and then DNA from the head of the virus can travel through the tail tube and enter the E. coli cell.
[citation needed] The lytic life cycle (from entering a bacterium to its destruction) takes approximately 30 minutes (at 37 °C).
These genetic experiments led to the finding of a unique linear order of mutational sites within the genes.
This result provided strong evidence for the key idea that the gene has a linear structure equivalent to a length of DNA with many sites that can independently mutate.
[citation needed] Penetration is also a value characteristic of phage-host infection that involves the injection of the phages genetic material inside the bacterium.
Nonenveloped viruses lyse the host cell which is characterized by viral proteins attacking the peptidoglycan or membrane.
DNA repair processes are also now recognized as playing critical roles in protecting against aging, cancer, and infertility.
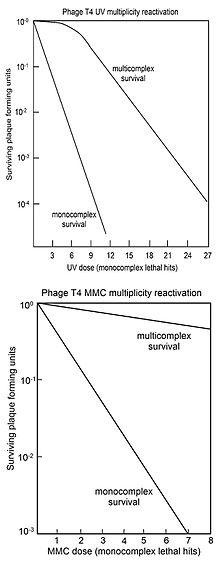
For comparison, the survival of virus plaque forming ability of singly infected cells (monocomplexes) is also plotted against dose of genome damaging agent.
The top figure shows the survival curves for virus T4 multicomplexes and monocomplexes with increasing dose of UV light.
Other virus T4 DNA damaging agents with shoulders in their multicomplex survival curves are X-rays[34][35] and ethyl methane sulfonate (EMS).
[36] The bottom figure shows the survival curves for inactivation of virus T4 by the DNA damaging agent mitomycin C (MMC).
In this case the survival curve for multicomplexes has no initial shoulder, suggesting that only the second recombinational repair process described above is active.
Similar multicomplex survival curves (without shoulders) were also obtained for the DNA damaging agents P32 decay, psoralen plus near-UV irradiation (PUVA), N-methyl-N'-nitro-N-nitrosoguanidine (MNNG), methyl methane sulfonate (MMS) and nitrous acid.
[citation needed] The specific time and place of T4 virus isolation remains unclear, though they were likely found in sewage or fecal material.
T4 and similar viruses were described in a paper by Thomas F. Anderson, Max Delbrück, and Milislav Demerec in November 1944.
The Luria–Delbrück experiment showed that bacteria, like other established model genetic organisms, have genes, and that these can spontaneously mutate to generate mutants that may then reproduce to form clonal lineages.
The phage group was an informal network of biologists centered on Max Delbrück that carried out basic research mainly on bacteriophage T4 and made numerous seminal contributions to microbial genetics and the origins of molecular biology in the mid-20th century.
In 1961, Sydney Brenner, an early member of the phage group, collaborated with Francis Crick, Leslie Barnett and Richard Watts-Tobin at the Cavendish Laboratory in Cambridge to perform genetic experiments that demonstrated the basic nature of the genetic code for proteins.
One noteworthy study used amber mutants defective in the gene encoding the major head protein of phage T4.
Other important scientists who worked with virus T4 include Michael Rossmann, Seymour Benzer, Bruce Alberts, Gisela Mosig,[48] Richard Lenski, and James Bull.