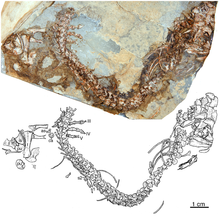
Caseidae
The most basal taxa were small insectivorous and omnivorous forms that lived mainly in the Upper Carboniferous and Lower Permian, such as Eocasea, Callibrachion, and Martensius.
The caseids experienced a significant evolutionary radiation at the end of the early Permian, becoming, with the captorhinid eureptiles, the dominant herbivores of terrestrial ecosystems in place of the edaphosaurids and diadectids.
With a fossil record spanning from the Late Carboniferous to the Middle Permian, caseids are one of the basal synapsids groups (formerly known as “pelycosaur”) having the largest stratigraphic range.
[nb 1] These last caseids still show a certain morphological diversity with medium to large herbivores (Ennatosaurus and Lalieudorhynchus), as well as small predatory or omnivorous forms such as Phreatophasma and possibly Eunotosaurus.
[1][2][3] They had a small head wider than high and with a forward-inclined snout, a very short neck, a long tail, robust forelimbs, and a body of variable proportions depending on their diet.
[4] Others with an omnivorous diet like Martensius had a barely enlarged rib cage, a more elongated skull, smaller nostrils, and a snout less inclined forward than in herbivorous caseids.
The postcranial skeleton indeed shows a spectacular increase in the volume of the rib cage, which becomes very wide and barrel-shaped, probably to accommodate a particularly developed intestine, necessary for the digestion of high-fiber rich plants.
In these forms, the skull has very large external nostrils and a very short facial region with a strong forward inclination of the end of the snout which clearly overhangs the dental row.
Instead of a dental battery, they had a massive tongue (as indicated by the presence of a highly developed hyoid apparatus found in Ennatosaurus and Euromycter) perhaps rough, with which they had to compress food against the palatal teeth.
These characteristics suggest that the initial strengthening of the forelimbs was probably related to a particular function such as digging, and that this trait was later exapted by more derived and larger caseids to support their weights of up to several hundred kilograms.
[nb 2] The most basal caseids like Eoasea, Callibrachion, and Martensius possessed the plesiomorphic condition of early amniotes with a phalangeal formula of 2-3-4-5-3 for manus and 2-3-4-5-4 for pes.
In the early 2000s, large footprints known as Brontopus giganteus from the Permian Lodève basin in southern France were considered to belong either to a caseid or dinocephalian therapsid.
[20] In 2019, Marchetti and colleagues, however, reinterpreted Chelichnus as a nomen dubium and a taphotaxon, this type of track showing false anatomical features generated by locomotion on sandy inclined paleosurface.
[21] In 2014, Eva Sacchi and colleagues described the ichnotaxon Dimetropus osageorum from about a hundred isolated footprints and several trackways from the early Permian Wellington Formation in Oklahoma.
The morphological study of these footprints and their comparison with tetrapod skeletons revealed that they probably belong to a large caseid comparable in size to Cotylorhynchus romeri (the latter from more recent strata).
[22] The study by Sacchi et al. also demonstrates that the ichnogenus Dimetropus exhibits great morphological variation and that its producers can be assigned to different zoological groups among non-therapsid synapsids and not just Sphenacodontidae as once thought.
[22] In 2021, Rafel Matamales-Andreu and colleagues assigned footprints found in the Lower Permian (Artinskian-Kungurian) Port des Canonge Formation in Mallorca to a caseid.
[23] During their evolutionary history, caseids shifted from faunivorous to herbivorous diet, a pattern that also occurred independently in other Carboniferous and Permian tetrapod groups such as Captorhinidae and Edaphosauridae.
Earliest and most basal caseids, such as the late Carboniferous Eocasea and the early Permian Callibrachion had an unexpanded rib cage and dentition composed of very small conical teeth suggesting an insectivorous diet.
These herbivorous caseids had spatulate teeth equipped with more or less numerous cuspules and a very enlarged and barrel-shaped rib cage which must have housed highly developed intestines necessary for the digestion of plants with low nutritional value.
Everett C. Olson in particular considered that the degree of ossification of the skeleton, the relatively short feet and hands, the massive claws, the limbs with very powerful extensor muscles, and the solid sacrum, strongly suggested a terrestrial lifestyle.
[25] This hypothesis is however disputed by Kenneth Angielczyk and Christian Kammerer as well as by Robert Reisz and colleagues based on paleontological and taphonomic data combined with the absence in these large caseids of morphological adaptations to an aquatic lifestyle.
According to Angielczyk and Kammerer, the low bone density of caseids identified by Lambertz et al. does not resemble that of semiaquatic animals, which tend to have a more strongly ossified skeleton to provide passive buoyancy control and increased stability against current and wave action.
Cotylorhynchus bone microstructure is more similar to what is seen in animals living in the open ocean, such as cetaceans and pinnipeds, which emphasize high maneuverability, rapid acceleration and hydrodynamic control of buoyancy.
[8] In 2022, Werneburg and colleagues proposed a somewhat different semiaquatic lifestyle, in which large caseids like Lalieudorhynchus (whose bone texture is even more osteoporotic than in Cotylorhynchus) would be ecological equivalents of modern hippos, passing part of their time in the water (being underwater walkers rather than swimming animals) but coming on dry land for food.
[26] Caseids first appear in the fossil record at the end of the Carboniferous and are already present in both North America and Europe with small predatory forms like Eocasea and Datheosaurus.
[27][26] Sedimentological and taphonomic data indicate that the majority of edaphosaurids favored humid habitats (their remains are found mainly in lacustrine and marshy deposits) while herbivorous caseids generally lived in drier biotopes where ponds and streams water were ephemeral.
[34] Caseid fossils have been found in the southern and central United States (Texas, Oklahoma, Kansas), France, Germany, Poland, Sardinia, and European Russia.
This geographical distribution corresponds to the paleoequatorial belt of Pangaea during the Carboniferous and the Permian, with the exception of the Russian localities which were located at the level of the 30th parallel north where a more temperate climate prevailed.
Lee and in 2021 by Asher J. Lichtig and Spencer G. Lucas, who reinterpreted the middle Permian taxon Eunotosaurus from South Africa as a small burrowing caseid.