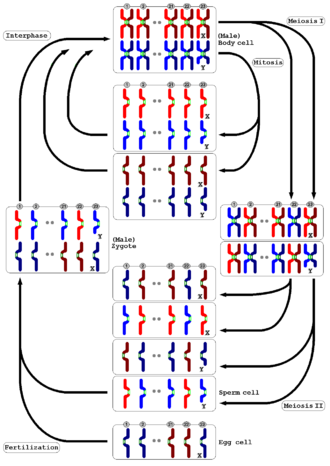
Meiosis
Errors in meiosis resulting in aneuploidy (an abnormal number of chromosomes) are the leading known cause of miscarriage and the most frequent genetic cause of developmental disabilities.
A subset of recombination events results in crossovers, which create physical links known as chiasmata (singular: chiasma, for the Greek letter Chi, Χ) between the homologous chromosomes.
In female animals, three of the four meiotic products are typically eliminated by extrusion into polar bodies, and only one cell develops to produce an ovum.
Female animals employ a slight variation on this pattern and produce one large ovum and three small polar bodies.
In some plants, fungi, and protists meiosis results in the formation of spores: haploid cells that can divide vegetatively without undergoing fertilization.
In 1911, the American geneticist Thomas Hunt Morgan detected crossovers in meiosis in the fruit fly Drosophila melanogaster, which helped to establish that genetic traits are transmitted on chromosomes.
[8]The spelling was changed to "meiosis" by Koernicke (1905) and by Pantel and De Sinety (1906) to follow the usual conventions for transliterating Greek.
Meiosis I and II are each divided into prophase, metaphase, anaphase, and telophase stages, similar in purpose to their analogous subphases in the mitotic cell cycle.
Meiosis II is an equational division analogous to mitosis, in which the sister chromatids are segregated, creating four haploid daughter cells (1n, 1c).
[21] This process generates single stranded DNA filaments coated by RAD51 and DMC1 which invade the homologous chromosomes, forming inter-axis bridges, and resulting in the pairing/co-alignment of homologues (to a distance of ~400 nm in mice).
[23] In this stage the homologous chromosomes become much more closely (~100 nm) and stably paired (a process called synapsis) mediated by the installation of the transverse and central elements of the synaptonemal complex.
In this stage homologous recombination, including chromosomal crossover (crossing over), is completed through the repair of the double strand breaks formed in leptotene.
Because the chromosomes cannot be distinguished in the synaptonemal complex, the actual act of crossing over is not perceivable through an ordinary light microscope, and chiasmata are not visible until the next stage.
During the diplotene stage, also known as diplonema, from Greek words meaning "two threads",[18]: 30 the synaptonemal complex disassembles and homologous chromosomes separate from one another a little.
Other than this observation, the rest of the stage closely resembles prometaphase of mitosis; the nucleoli disappear, the nuclear membrane disintegrates into vesicles, and the meiotic spindle begins to form.
[32] This is followed by anaphase II, in which the remaining centromeric cohesin, not protected by Shugoshin anymore, is cleaved, allowing the sister chromatids to segregate.
As one example, Giardia intestinalis, a common intestinal parasite, was previously considered to have descended from a lineage that predated the emergence of meiosis and sex.
[40] Although synaptonemal complexes are present during the pachytene stage of meiosis in B. mori, crossing-over homologous recombination is absent between the paired chromosomes.
The arrest of ooctyes at the four genome copy stage was proposed to provide the informational redundancy needed to repair damage in the DNA of the germline.
[43] DNA repair capability appears to be a key quality control mechanism in the female germ line and a critical determinant of fertility.
[47] Volvox carteri, a haploid multicellular, facultatively sexual green algae, can be induced by heat shock to reproduce by meiotic sex.
The organism's diploid germ-line stem cells undergo meiosis to make haploid gametes (the spermatozoa in males and ova in females), which fertilize to form the zygote.
[citation needed] In the haplodiplontic life cycle (with sporic or intermediate meiosis), the living organism alternates between haploid and diploid states.
When they germinate, these spores undergo repeated cell division by mitosis, developing into a multicellular haploid gametophyte generation, which then produces gametes directly (i.e. without further meiosis).
In both animals and plants, the final stage is for the gametes to fuse to form a zygote in which the original number of chromosomes is restored.
Maturing oocytes are arrested in prophase I of meiosis I and lie dormant within a protective shell of somatic cells called the follicle.
The breakdown of the germinal vesicle, condensation of chromosomes, and assembly of the bipolar metaphase I spindle are all clear indications that meiosis has resumed.
[71] Meiotic arrest requires inhibitory phosphorylation of CDK1 at amino acid residues Thr-14 and Tyr-15 by MYT1 and WEE1[52] as well as regulation of cyclin B levels facilitated by the anaphase-promoting complex (APC).
[52] The protein kinase p90 (RSK) is one critical target of MAPK and may help block entry into S-phase between meiosis I and II by reactivating CDK1.
[75] IME2/SME1 enables sporulation when nitrogen is present, supports recombination in a/α cells expressing RME1, an inhibitor of meiosis, and encodes a protein kinase homolog.