GroEL
In eukaryotes the organellar proteins Hsp60 and Hsp10 are structurally and functionally nearly identical to GroEL and GroES, respectively, due to their endosymbiotic origin.
It may facilitate the correct folding of imported proteins, and may also prevent misfolding and promote the refolding and proper assembly of unfolded polypeptides generated under stress conditions in the mitochondrial matrix.
In addition to its role as a heat shock protein, HSP60 functions as a chaperonin to assist in folding linear amino acid chains into their respective three-dimensional structure.
This protein presence provides evidence for the evolutionary relationship of the development of the mitochondria and the chloroplast by means of endosymbiosis.
[6] Under normal physiological conditions, HSP60 is a 60 kilodalton oligomer composed of monomers that form a complex arranged as two stacked heptameric rings.
[8] This double ring structure forms a large central cavity in which the unfolded protein binds via hydrophobic interactions.
[10] Recent studies have begun to suggest that in addition to its typical location in the mitochondria, HSP60 can also be found in the cytoplasm under normal physiological conditions.
[11] The intermediate domain induces a conformational change when ATP is bound allowing for an alternation between the hydrophilic and hydrophobic substrate binding sites.
When activated by ATP, the intermediate domain undergoes a conformational change that exposes the hydrophilic region.
The N-terminal contains a pre-sequence of hydroxylated amino acids, namely arginine, lysine, serine, and threonine, which serve as directors for the importation of the protein into the mitochondria.
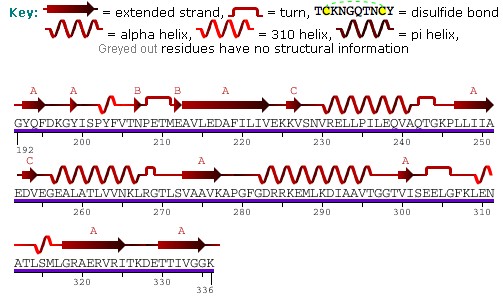
[6] The predicted structure of HSP60 includes several vertical sine waves, alpha helices, beta sheets, and 90 degree turns.
The hydrophobic portion HSP60 is responsible for maintaining the unfolded conformation of the protein for transmembrane transport.
[14] Studies have shown how HSP60 binds to incoming proteins and induces conformational and structural changes.
In addition to its critical role in protein folding, HSP60 is involved in the replication and transmission of mitochondrial DNA.
In extensive studies of HSP60 activity in Saccharomyces cerevisiae, scientists have proposed that HSP60 binds preferentially to the single stranded template DNA strand in a tetradecamer like complex [15] This tetradecamer complex interacts with other transcriptional elements to serve as a regulatory mechanism for the replication and transmission of mitochondrial DNA.
HSP60 monomers form two heptameric rings that bind to the surface of linear proteins and catalyze their folding in an ATP dependent process.
[9] The necessity of preexisting HSP60 in order to synthesize additional HSP60 molecules supports the endosymbiotic theory of the origin of mitochondria.
Since they are very similar in sequence, bacterial HSP60 wouldn’t be expected to cause a large immune response in humans.
The heat shock response is a homeostatic mechanism that protects a cell from damage by upregulating the expression of genes that code for HSP60.
[20] The upregulation of HSP60 production allows for the maintenance of other cellular processes occurring in the cell, especially during stressful times.
Researchers concluded that the heat shock signal pathway serves as “the basic mechanism of defense against neurotoxicity elicited by free radical oxygen and nitrogen species produced in aging and neurodegenerative disorders”.
[21] Several studies have shown that HSP60 and other heat shock proteins are necessary for cellular survival under toxic or stressful circumstances.
[22] Human Hsp60, the product of the HSPD1 gene, is a Group I mitochondrial chaperonin, phylogenetically related to bacterial GroEL.
This chaperone was also classically regarded as an intracellular protein like Hsp60, but in the last few years considerable evidences showed its pericellular and extracellular residence HSP60 has been shown to influence apoptosis in tumor cells which seems to be associated with a change in expression levels.
[23][24] As well as influencing apoptosis, HSP60 changes in expression level have been shown to be “useful new biomarkers for diagnostic and prognostic purposes.” [23] According to Lebret et al., a loss of HSP60 expression “indicates a poor prognosis and the risk of developing tumor infiltration” specifically with bladder carcinomas, but that does not necessarily hold true for other types of cancers.
[25] All this research indicates that it may be possible for HSP60 expression to be used in predicting survival for certain types of cancer and therefore may be able to identify patients who could benefit from certain treatments.
Binding of GroES to the open cavity of the chaperonin induces the individual subunits of the chaperonin to rotate such that the hydrophobic substrate binding site is removed from the interior of the cavity, causing the substrate protein to be ejected from the rim into the now largely hydrophilic chamber.
The geometry of GroEL requires that the polar structures lead, and they envelop the non-polar core as it emerges from the trans side.
Many globular proteins won't bind to the apical domain because their hydrophobic parts are clustered inside, away from the aqueous medium since this is the thermodynamically optimal conformation.
This effect, multiplied through the cooperative flexing of all monomers, increases the equatorial diameter of the GroEL cage.

{kind=link}