Knockout rat
[5] Additional developments with zinc finger nuclease technology in 2009 led to the first knockout rat with targeted, germline-transmitted mutations.
[6] Knockout rat disease models for Parkinson's, Alzheimer's, hypertension, and diabetes using zinc-finger nuclease technology are being commercialized by SAGE Labs.
It is widely believed that the rat is a better model than the mouse for human cardiovascular disease, diabetes, arthritis, and many autoimmune, neurological, behavioral, and addiction disorders.
[12] Their larger size makes rats more conducive to study by instrumentation, and also facilitates manipulation such as blood sampling, nerve conduction, and performing surgeries.
ENU transfers its ethyl group to oxygen or nitrogen radicals in DNA, resulting in mis-pairing and base pair substitution.
ENU mutagenesis creates a high frequency of random mutations, with approximately one base pair change in any given gene in every 200-700 gametes.
ENU and cloning methods are simply inefficient for creating and mapping gene knockouts in rats for the creation of new models of human disease.
Although some companies are still pursuing this strategy, the Medical College of Wisconsin has switched to a more efficient and commercially viable method using mobile DNA and CompoZr ZFN technology.
Double strand breaks are important for site-specific mutagenesis in that they stimulate the cell's natural DNA-repair processes, namely homologous recombination and non-homologous end joining.
This platform technology meets all of the criteria for a successful gene knockout approach in mammals by permitting random mutagenesis directly in the germ cells (sperm and oocytes) of mammalian model organisms, including rats.
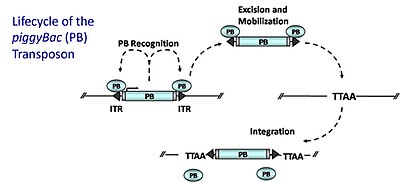
PB possesses several ideal properties: (1) it preferentially inserts within genes (50 to 67% of insertions hit genes) (2) it exhibits no local hopping (widespread genomic coverage) (3) it is not sensitive to over-production inhibition in which elevated levels of the transposase cause decreased transposition 4) it excises cleanly from a donor site, leaving no “footprint,” unlike Sleeping Beauty.
[28][29][30][31][32][33][34] There are several advantages that make SB a highly attractive mutagen geared toward gene discovery: 1) it has little bias for inserting within particular genomic regions or within specific recognition sequences, 2) de novo insertions of the transposon provide a “tagged” sequence marker for rapid identification of the specific mutation by simple PCR cloning methods, 3) in vivo SB insertional mutagenesis allows multiple mutations to be quickly and easily generated in a single animal, and in a single tissue, such as an adenomatous polyp.
Retrotransposons, such as LINEs (long interspersed nuclear elements), mobilize via a “copy and paste” mechanism and are abundant in many eukaryotic species.
These proteins exhibit an overwhelming specificity for binding to and acting on the transcript that encodes them, enabling near exclusive mobilization of the parental L1 RNA.
Using the RT activity of the ORF2 protein, the transcribed L1 RNA is copied into DNA by a process termed target primed reverse transcription (TPRT),[35] and integrated into the genome.
The nature of retrotransposition endows the L1 with some unique advantages; L1 retrotransposons have an essentially unlimited supply of the insertional mutagen since it is continually transcribed from a promoter, which would be useful for applications where large numbers of mutations are needed in a single cell.