Lake metabolism
[2] Ecosystem respiration in excess of gross primary production indicates the lake receives organic material from the surrounding catchment, such as through stream or groundwater inflows or litterfall.
Cyclical changes on a daily scale occur in most lakes on Earth because sunlight is available for photosynthesis and production of new carbon only for a portion of the day.
The following sections highlight the most common ways to estimate lake metabolism across a variety of temporal and spatial scales and go over some of the assumptions of each of these methods.
Abiotic changes in dissolved gases can dominate changes of dissolved gases if the lake has a low metabolic rate (e.g. oligotrophic lake, cloudy day), or if there is a large event that causes abiotic factors to exceed biotic (e.g. wind event causing mixing and entrainment of low-oxygenated water).
Atmospheric gas exchange (F) is rarely directly measured and typically modeled by estimating lake surface turbulence from wind-driven and convective mixing.
Analogous to the light and dark bottle methods described above, lake sediment cores can be collected and changes in dissolved oxygen or carbon fixation can be used to estimate rates of primary productivity and respiration.
For example, fast-growing cladocerans have a much lower nitrogen to phosphorus ratio (N:P) than copepods, mostly due to the high amount of phosphorus-rich RNA in their cells used for rapid growth.
In addition to controlling primary productivity, sunlight can also influence rates of respiration by partially oxidizing organic matter which can make it easier for bacteria to break down and convert into carbon dioxide.
During algal blooms, elevated dissolved CO2 ensures that CO2 is not a limiting resource for growth since rapid increases in production deplete CO2 and raise pH.
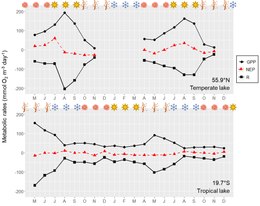
Metabolic rates in lakes and reservoirs are controlled by many environmental factors, such as light and nutrient availability, temperature, and water column mixing regimes.
Thus, littoral zones are more susceptible to changes in temperature, inputs of nutrients and organic matter from the landscape and river inflows, wind shear mixing and wave action, shading from terrestrial vegetation, and resuspension of the sediments (Figure 1).
Additionally, littoral zones usually have greater habitat complexity due to the presence of macrophytes, which serve as shelter, nursery, and feeding place for many organisms.
Relatively small forested lakes are more shaded and protected from wind exposure and also receive high amounts of allochthonous organic matter.
The high inputs of allochthonous organic matter (produced outside the lake) stimulate heterotrophic communities, such as bacteria, zooplankton, and fish, enhancing whole-lake respiration rates.
As the sun rises, there is a rapid increase in primary production in the lake, often making it autotrophic (NEP > 0) and reducing dissolved CO2 that was produced from carbon mineralization that occurred during the night.
[51] These weather variations also cause short-term variability in mixed layer depth, which in turn affects nutrients, organic matter, and light availability, as well as vertical and horizontal gas exchanges.
Thus the effects of short-term variability in mixed layer depth on gross primary production (GPP) will depend on which are the limiting factors on each lake at a given period.
Thus a deeper mixing layer could either increase or decrease GPP rates depending on the balance between nutrient and light limitation of photosynthesis (Figure).
Responses in metabolic rates are as dynamic as the physical and chemical processes occurring in the lake, but changes in algal biomass are less variable, involving growth and loss over longer periods.
Moreover, during seasons with higher light levels photodegradation of organic matter is more pronounced, which stimulates microbial degradation, enhancing heterotrophic respiration rates.
Most of the lakes in the world freeze during the winter,[53] a low-irradiance period, in which ice and snow cover limit light penetration in the water column.
Regardless of the high GPP in these systems, ER rates are also enhanced by the increased availability of organic matter stocks from sediments and deeper waters.
On the other hand, lower precipitation also affects limnological conditions by reducing the water level and, thereby, increasing the concentration of nutrients and chlorophyll, as well as changing the thermal stability of aquatic environments.
Lakes frequently exposed to strong storms, such as the typhoon areas in the Northwest Pacific Ocean, receive intense rainfall events that can last for a few days.
While lower precipitation associated with high evaporation rates also affects limnological conditions by reducing the water level and thereby increasing the concentration of nutrients and chlorophyll, as well as changing the thermal stability of aquatic environments.
Thus, the effects of differences in precipitation and temperature between years in metabolic rates will depend on the intensity and duration of these changes, and also in which factors are limiting GPP and ER in each water body.
This will depend if the terrestrial inputs will be promptly available for the primary producers and heterotrophic communities or if it is going to enter the lake through deeper waters, in which metabolic processes are very low or non-existent.
On the other hand, drier years can also have enhanced GPP and ER rates if it is accompanied by lower water levels, which would lead to higher nutrients and organic matter concentrations.
In lakes with a photic zone extending deeper than the mixed layer, metabolic rates will be higher in intermediated depths, coinciding with the deep chlorophyll maxima.
Williamson and collaborators,[49] for example, found that, in a hyper-eutrophic reservoir in North America, the relative spatial variability in GPP and ER rates were higher in a dry year compared to a wet one.