Mushroom bodies
The mushroom bodies or corpora pedunculata are a pair of structures in the brain of arthropods, including insects and crustaceans,[2] and some annelids (notably the ragworm Platynereis dumerilii).
In most insects, the mushroom bodies and the lateral horn are the two higher brain regions that receive olfactory information from the antennal lobe via projection neurons.
Most of our current knowledge of mushroom bodies comes from studies of a few species of insect, especially the cockroach Periplaneta americana, the honey bee Apis mellifera,[7] the locust and the fruit fly Drosophila melanogaster.
They are mainly composed of the long, densely packed nerve fibres of the Kenyon cells, the intrinsic neurons of the mushroom bodies.
[10] In Hymenoptera in particular, subregions of the mushroom body neuropil are specialized to receive olfactory, visual, or both types of sensory input.
Research implies that mushroom bodies generally act as a sort of coincidence detector, integrating multi-modal inputs[4] and creating novel associations, thus suggesting their role in learning and memory.
[18] Experiments in locusts have shown that Kenyon cells have their activity synchronized to 20-Hz neural oscillations and are particularly responsive to projection neuron spikes at specific phases of the oscillatory cycle.
A topic of current research is which of these substructures in the mushroom body are involved in each phase and process of learning and memory.
[24] Recent studies combining odor conditioning and cellular imaging have identified six memory traces that coincide with molecular changes in the Drosophila olfactory system.
[27] The intermediate term memory trace is dependent on expression of the amn gene located in dorsal paired medial neurons.
An increase in calcium influx and synaptic release that innervates the mushroom bodies becomes detectable approximately 30 minutes after pairing of electric shock with an odor, and persists for at least an hour.
[28] Both long-term memory traces that have been mapped depend on activity and protein synthesis of CREB and CaMKII, and only exist after spaced conditioning.
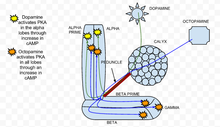
Therefore, rut AC in mushroom body neurons works as a coincidence detector with dopamine and octopamine functioning bidirectionally to affect cAMP levels.
[33] When calcium enters a cell and binds with calmodulin, it stimulates adenylate cyclase (AC), which is encoded by the rutabaga gene (rut).
[34] When dopamine, an aversive olfactory stimulant, is applied it activates PKA specifically in the vertical mushroom body lobes.
[34] In the rut mutant, a genotype in which the rutabaga is abolished, the responses to both dopamine and octopamine were greatly reduced and close to experimental noise.
Acetylcholine, which represents the conditioned stimulus, leads to a strong increase in PKA activation compared to stimulation with dopamine or octopamine alone.