Natural competence
Artificial competence arises when cells in laboratory cultures are treated to make them transiently permeable to DNA.
[1][2] In 1944 Oswald Avery, Colin MacLeod, and Maclyn McCarty demonstrated that this 'transforming factor' was pure DNA.
Since then, natural competence has been studied in a number of different bacteria, particularly Bacillus subtilis, Streptococcus pneumoniae, Neisseria gonorrhoeae, Haemophilus influenzae and members of the Acinetobacter genus.
[5] In laboratory cultures, natural competence is usually tightly regulated and often triggered by nutritional shortages or adverse conditions.
Most naturally competent bacteria are thought to take up all DNA molecules with roughly equal efficiencies.
[12] However, since bacteria tend to grow in clones, the DNA available for transformation would generally have the same genotype as that of the recipient cells.
In principle the cost of nuclease production and the uncertainty of nucleotide recovery must be balanced against the energy needed to synthesize the uptake machinery and to pull DNA in.
Other important factors are the likelihoods that nucleases and competent cells will encounter DNA molecules, the relative inefficiencies of nucleotide uptake from the environment and from the periplasm (where one strand is degraded by competent cells), and the advantage of producing ready-to-use nucleotide monophosphates from the other strand in the cytoplasm.
Some bacteria induce competence under such stress conditions, supporting the hypothesis that transformation helps DNA repair.
[19][20][1] Thus, a strong short-term selective advantage for natural competence and transformation would be its ability to promote homologous recombinational DNA repair under conditions of stress.
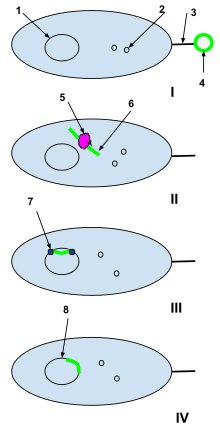
1-Bacterial cell DNA
2-Bacterial cell plasmids
3-Sex pili
4-Plasmid of foreign DNA from a dead cell
5-Bacterial cell restriction enzyme
6-Unwound foreign plasmid
7-DNA ligase
I: A plasmid of foreign DNA from a dead cell is intercepted by the sex pili of a naturally competent bacterial cell.
II: The foreign plasmid is transduced through the sex pili into the bacterial cell, where it is processed by bacterial cell restriction enzymes. The restriction enzymes break the foreign plasmid into a strand of nucleotides that can be added to the bacterial DNA.
III: DNA ligase integrates the foreign nucleotides into the bacterial cell DNA.
IV: Recombination is complete and the foreign DNA has integrated into the original bacterial cell's DNA and will continue to be a part of it when the bacterial cell replicates next.