RNA editing
[1][2][3][4] RNA editing has been observed in some tRNA, rRNA, mRNA, or miRNA molecules of eukaryotes and their viruses, archaea, and prokaryotes.
[7] RNA-editing processes show great molecular diversity, and some appear to be evolutionarily recent acquisitions that arose independently.
The diversity of RNA editing phenomena includes nucleobase modifications such as cytidine (C) to uridine (U) and adenosine (A) to inosine (I) deaminations, as well as non-template nucleotide additions and insertions.
[21] Moreover, mass spectrometry allows the investigation of modification dynamics by labelling RNA molecules with stable (non-radioactive) heavy isotopes in vivo.
This method, called NAIL-MS (nucleic acid isotope labelling coupled mass spectrometry), enables a variety of approaches to investigate RNA modification dynamics.
They ensure the correct maturation and function of the mRNA, but also at the same time act as part of cell's immune system.
[27] For example, m6A has been predicted to affect protein translation and localization,[1][2][3] mRNA stability,[28] alternative polyA choice [14] and stem cell pluripotency.
[29] Pseudouridylation of nonsense codons suppresses translation termination both in vitro and in vivo, suggesting that RNA modification may provide a new way to expand the genetic code.
[1] For example, genetic mutations in pseudouridine synthases cause mitochondrial myopathy, sideroblastic anemia (MLASA) [32] and dyskeratosis congenital.
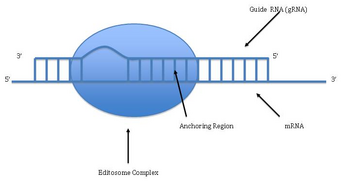
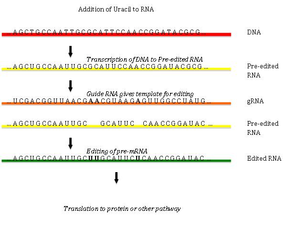
Pan-editing starts with the base-pairing of the unedited primary transcript with a guide RNA (gRNA), which contains complementary sequences to the regions around the insertion/deletion points.
The newly formed double-stranded region is then enveloped by an editosome, a large multi-protein complex that catalyzes the editing.
The mechanism of the editosome involves an endonucleolytic cut at the mismatch point between the guide RNA and the unedited transcript.
In the intestines, the mRNA has a CAA sequence edited to be UAA, a stop codon, thus producing the shorter B48 form.
Studies have shown that I-RNA (RNA with many repeats of the I-U base pair) recruits methylases that are involved in the formation of heterochromatin and that this chemical modification heavily interferes with miRNA target sites.
Newer studies, however, have weakened this correlation by showing that inosines can also be decoded by the ribosome (although in a lesser extent) as adenosines or uracils.
It has been shown in previous studies that the only types of RNA editing seen in the plants' mitochondria and plastids are conversion of C-to-U and U-to-C (very rare).
While the specific positions for C to U RNA editing events have been fairly well studied in both the mitochondrion and plastid,[80] the identity and organization of all proteins comprising the editosome have yet to be established.
Members of the expansive PPR protein family have been shown to function as trans-acting factors for RNA sequence recognition.
[82] An enzyme responsible for the trans- or deamination of the RNA transcript remains elusive, though it has been proposed that the PPR proteins may serve this function as well.
Therefore, it is highly probable that polypeptides synthesized from unedited RNAs would not function properly and hinder the activity of both mitochondria and plastids.
Viruses are known to utilize the RNA modifications in different parts of their infection cycle from immune evasion to protein translation enhancement.
[84][85] Viral RNAs are transcribed by a virus-encoded RNA-dependent RNA polymerase, which is prone to pausing and "stuttering" at certain nucleotide combinations.
Additionally, the RNA modifications are shown to have both positive and negative effects on the replication and translation efficiency depending on the virus.
For example, Courtney et al.[12] showed that an RNA modification called 5-methylcytosine is added to the viral mRNA in infected host cells in order to enhance the protein translation of HIV-1 virus.
However, this active site is widened by amino acid changes in the corresponding human analog genes, APOBEC1 and ADAR, allowing deamination.
[88][89] The gRNA-mediated pan-editing in trypanosome mitochondria, involving templated insertion of U residues, is an entirely different biochemical reaction.
[43][90] But the specificity of nucleotide insertion via the interaction between the gRNA and mRNA is similar to the tRNA editing processes in the animal and Acanthamoeba mitochondria.
[91] Eukaryotic ribose methylation of rRNAs by guide RNA molecules is a similar form of modification.
A more plausible alternative for the evolutionary origins of this system is through constructive neutral evolution, where the order of steps is reversed, with the gratuitous capacity for editing preceding the "defect".
Therefore, the changes led >25% correction of the targeted stop codon with read through to a downstream luciferase reporter sequence.