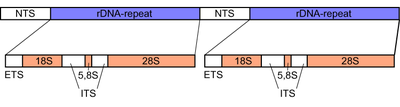
Ribosomal DNA
[6] Similar to the structure of prokaryotes, the 5S rRNA is appended to the rDNA cluster in the Saccharomycetes (Hemiascomycetes)[6] such as Saccharomyces cerevisiae.
[1] The human genome contains a total of 560 copies[4] of the rDNA transcriptional unit, spread across five chromosomes with nucleolus organizer regions.
[7] In ciliates, the presence of a generative micronucleus next to the vegetative macronucleus allows for the reduction of rDNA genes in the germline.
During macronucleus formation, the regions containing the rDNA gene clusters are amplified, dramatically increasing the amount of available templates for transcription up to several thousand copies.
In some ciliate genera, such as Tetrahymena or the Hypotrich genus Oxytricha,[8] extensive fragmentation of the amplified DNA leads to the formation of microchromosomes, centered on the rDNA transcriptional unit.
[6] However, the mechanism of concerted evolution is imperfect, such that polymorphisms between repeats within an individual can occur at significant levels and may confound phylogenetic analyses for closely related organisms.
[11] They could occur by slippage of the newly synthesized strand during DNA replication or by gene conversion.
[11] The rDNA transcription tracts have low rate of polymorphism among species, which allows interspecific comparison to elucidate phylogenetic relationship using only a few specimens.
[12] Conserved sequences at coding regions of rDNA allow comparisons of remote species, even between yeast and human.
As a result, this DNA can provide phylogenetic information of species belonging to wide systematic levels.
Also, any damage or disruption to the enzymes that protect the tandem repeats of the rDNA, can result in lower synthesis of ribosomes, which also lead to other defects in the cell.