Regulatory T cell
[3] Mouse models have suggested that modulation of Treg cells can treat autoimmune disease and cancer and can facilitate organ transplantation[4] and wound healing.
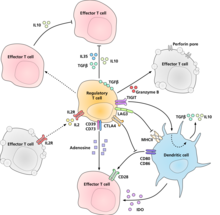
[9] Regulatory T cells are involved in shutting down immune responses after they have successfully eliminated invading organisms, and also in preventing autoimmunity.
The selection of Tregs occurs on radio-resistant hematopoietically derived MHC class II-expressing cells in the medulla or Hassall's corpuscles in the thymus.
Tregs expressing low amounts of CD25, GITR and PD-1 limit the development of colitis by promoting the conversion of conventional CD4+ T cells into pTreg.
Tregs highly expressing CD25, GITR and PD-1 are more self-reactive and control lymphoproliferation in peripheral lymph nodes - they may have the ability to protect against autoimmune disorders.
[18] Recirculating Tregs in the thymus express high levels of the high-affinity IL-2 receptor α chain (CD25), encoded by the Il2ra gene, which gathers IL-2 from thymic medulla and decreases its concentration.
When self/non-self discrimination fails, the immune system destroys cells and tissues of the body and as a result causes autoimmune diseases.
[37] Natural T regulatory lymphocytes (tTregs, nTregs) are characterized by continuous expression of FoxP3 and T cell receptor (TCR) with relatively high autoaffinity.
These cells are predominantly found in the body in the bloodstream or lymph nodes and serve mainly to confer tolerance to autoantigens.
[37] The main features that differentiate tTreg and iTreg cells include Helios and Neuropilin-1, the presence of which suggests origin in the thymus.
[40][41] The intestinal environment can lead to induced regulatory T cells with TGF-beta and retinoic acid,[42] some of which express the lectin-like receptor CD161 and are specialized to maintain barrier integrity by accelerating wound healing.
[48] Hp-TGM-induced human FOXP3+ regulatory T cells were stable in the presence of inflammation and had increased levels of CD25, CTLA4 and decreased methylation in the FOXP3 Treg-Specific demethylated region compared to TGF-β-induced Tregs.
In the intestinal lamina propria, 20-30% of Foxp3+ T regulatory cells expressing RORyt are found and this high proportion is strongly dependent on the presence of a complex gut microbiome.
In germ-free (GF) mice, the population of RORγt+ T regulatory cells is strongly reduced, whereas recolonization by the specific pathogen-free (SPF) microbiota restores normal numbers of these lymphocytes in the gut.
The mechanism by which the gut microbiota induces the formation of RORγt+ Treg cells involves the production of short-chain fatty acids (SCFAs), on which this induction is dependent.
Particularly crucial is a time period of gradual transition from relying solely on maternal milk to incorporating solid food, between 15 and 20 days of age, when a large number of microbial antigens is introduced and commensal microbiota are settling in the intestine.
During this time, protective RORγt+ Treg cells are induced by the microbial antigens and normal intestinal homeostasis is sustained by induction of tolerance to commensal microbiota.
This induction is possibly regulated by stimulation of Aryl hydrocarbon receptor by metabolites produced by commensal bacteria using tryptophan as an energy source.
[58] In tumors of mice with conditional knockout of RORγt+ Tregs was confirmed downregulation of IL-6, reduction of IL-6 expressing CD11c+ dendritic cells and overexpression of CTLA-4.
While the immunosuppressive function of regulatory T cells prevents the development of autoimmune disease, it is not desirable during immune responses to infectious microorganisms.
Experimental evidence from mouse models suggests that some pathogens may have evolved to manipulate regulatory T cells to immunosuppress the host and so potentiate their own survival.
There is some evidence that Treg cells may be dysfunctional and driving neuroinflammation in amyotrophic lateral sclerosis due to lower expression of FOXP3.
[65] Ex vivo expansion of Treg cells for subsequent autologous transplant is currently being investigated after promising results were obtained in a phase I clinical trial.
[72] These opposite effects indicate that Tr cells' role in the development of cancer is highly dependent on both type and location of the tumor.
It has been shown that Tregs can be detected using an activation-induced marker assay by expression of CD39[82] in combination with co-expression of CD25 and OX40(CD134) which define antigen-specific cells following 24-48h stimulation with antigen.
[83][84] In addition to the search for novel protein markers, a different method to analyze and monitor Treg cells more accurately has been described in the literature.
[87] Regulatory T cell epitopes ('Tregitopes') were discovered in 2008 and consist of linear sequences of amino acids contained within monoclonal antibodies and immunoglobulin G (IgG).
[88][89][90] Potential applications of regulatory T cell epitopes have been hypothesised: tolerisation to transplants, protein drugs, blood transfer therapies, and type I diabetes as well as reduction of immune response for the treatment of allergies.
This disease provides the most striking evidence that regulatory T cells play a critical role in maintaining normal immune system function.
Humans with mutations in FOXP3 develop a severe and rapidly fatal autoimmune disorder known as Immune dysregulation, Polyendocrinopathy, Enteropathy X-linked (IPEX) syndrome.