Sodium channel
Even though the resting and inactive states would not allow the ions to flow through the channels the difference exists with respect to their structural conformation.
When accessory proteins assemble with α subunits, the resulting complex can display altered voltage dependence and cellular localization.
[5] When stimulated by a change in transmembrane voltage, this segment moves toward the extracellular side of the cell membrane, allowing the channel to become permeable to ions.
This DIII-IV linker is responsible for wedging the pore gate shut after channel opening, inactivating it.
The inactivation gate can be thought of as a "plug" tethered to domains III and IV of the channel's intracellular alpha subunit.
This refractory period eliminates the possibility of an action potential moving in the opposite direction back towards the soma.
The refractory period of each channel is therefore vital in propagating the action potential unidirectionally down an axon for proper communication between neurons.
Mutations that interfere with Na+ channel inactivation can contribute to cardiovascular diseases or epileptic seizures by window currents, which can cause muscle and/or nerve cells to become over-excited.
The cations flow into a more constricted part of the pore that is 0.3 by 0.5 nm wide, which is just large enough to allow a single Na+ ion with a water molecule associated to pass through.
Ions of different sizes also cannot interact as well with the negatively charged glutamic acid residues that line the pore.
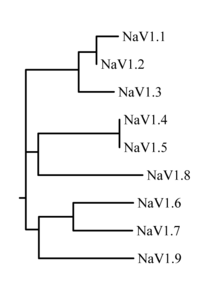
The family of sodium channels has 9 known members, with amino acid identity >50% in the trans-membrane segments and extracellular loop regions.
[12] The probable evolutionary relationship between these channels, based on the similarity of their amino acid sequences, is shown in figure 1.
Gastrointestinal: Irritable bowel syndrome;[17] Sodium channel beta subunits are type 1 transmembrane glycoproteins with an extracellular N-terminus and a cytoplasmic C-terminus.
[22] Sodium channels are more likely to stay open at the subthreshold membrane potential when interacting with beta toxins, which in turn induces an immediate sensation of pain.
[10][24][25] Voltage-gated sodium channels also assemble with a variety of other proteins, such as FHF proteins (Fibroblast growth factor Homologous Factor), calmodulin, cytoskeleton or regulatory kinases,[26][10][27][28][29] which form a complex with sodium channels, influencing its expression and/or function.
The ability of these channels to assume a closed-inactivated state causes the refractory period and is critical for the propagation of action potentials down an axon.
Ligand-gated sodium channels, on the other hand, create the change in the membrane potential in the first place, in response to the binding of a ligand to it.
Instead, they are always open or "leaking" a small background current to regulate the resting membrane potential of a neuron.
[35] In most animals, a single gene encodes the NALCN (sodium leak channel, nonselective) protein.
[35][42][41][40] FAM155A helps protein folding in the endoplasmic reticulum, chaperones transport to the axon, and contributes to membrane stability.
Mutations to NALCN lead to severe disruptions to respiratory rhythm in mice[39] and altered circadian locomotion in flies.
[49] Changes in blood and tissue pH accompany physiological and pathophysiological conditions such as exercise, cardiac ischemia, ischemic stroke, and cocaine ingestion.
These conditions are known to trigger the symptoms of electrical diseases in patients carrying sodium channel mutations.
These effects are shared with disease-causing mutants in neuronal, skeletal muscle, and cardiac tissue and may be compounded in mutants that impart greater proton sensitivity to sodium channels, suggesting a role of protons in triggering acute symptoms of electrical disease.
[51] The sodium channel selectivity filter is composed of a single residue in each of the four pore-loops of the four functional domains.
This has been suggested to be a protective mechanism against potential over- or under-excitability in skeletal muscles, as blood pH levels are highly susceptible to change during movement.