AMP-activated protein kinase
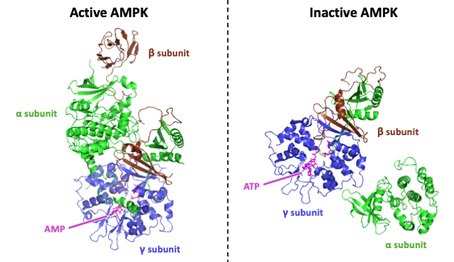
[4][5] Specifically, the γ subunit includes four particular Cystathionine-β-synthase (CBS) domains, giving AMPK its ability to sensitively detect shifts in the AMP/ATP ratio.
Upon KD dissociation, the active loop (AL) of the α subunit which contains the critical phosphorylated Thr residue is fully exposed to upstream phosphatases.
[16] AMPK stimulates glucose uptake in skeletal muscle by phosphorylating Rab-GTPase-activating protein TBC1D1, which ultimately induces fusion of GLUT4 vesicles with the plasma membrane.
[16] AMPK also appears to stimulate mitochondrial biogenesis by regulating PGC-1α which in turn promotes gene transcription in mitochondria.
[26][27] Additionally, recent discoveries can conceivably suggest a direct AMPK role in increasing blood supply to exercised/trained muscle cells by stimulating and stabilizing both vasculogenesis and angiogenesis.
During a single acute exercise bout, AMPK allows the contracting muscle cells to adapt to the energy challenges by increasing expression of hexokinase II,[23] translocation of GLUT4 to the plasma membrane,[29][30][31][32] for glucose uptake, and by stimulating glycolysis.
[33] If bouts of exercise continue through a long-term training regimen, AMPK and other signals will facilitate contracting muscle adaptations by escorting muscle cell activity to a metabolic transition resulting in a fatty-acid oxidation approach to ATP generation as opposed to a glycolytic approach.
[26][23][25][34][35][36][37] Mutations in the skeletal muscle calcium release channel (RYR1) underlies a life- threatening response to heat in patients with malignant hyperthermia susceptibility (MHS).
Upon acute exposure to heat, these mutations cause uncontrolled Ca2+ release from the sarcoplasmic reticulum, leading to sustained muscle contractures, severe hyperthermia, and sudden death.
[45] Their study compared the response to exercise training of several proteins and enzymes in wild type and AMPKα2 knockout mice.
These findings are supported by another study also showing no difference in mitochondrial adaptations to exercise training between wild type and knockout mice.
[46] The C. elegans homologue of AMPK, aak-2, has been shown by Michael Ristow and colleagues to be required for extension of life span in states of glucose restriction mediating a process named mitohormesis.
One of the key pathways in AMPK's regulation of fatty acid oxidation is the phosphorylation and inactivation of acetyl-CoA carboxylase.
[29] HMGR converts 3-hydroxy-3-methylglutaryl-CoA, which is made from acetyl-CoA, into mevalonic acid, which then travels down several more metabolic steps to become cholesterol.
[48][23][34] Chronic AICAR injections, simulating some of the effects of endurance training, also increase the total amount of GLUT-4 protein in the muscle cell.
[36] With chronic injections of AICAR, total protein content of hexokinase II increases in rat skeletal muscle.
[43] AICAR stimulation of AMPK increases cytochrome c and δ-aminolevulinate synthase (ALAS), a rate-limiting enzyme involved in the production of heme.
[50] AMPK is required for increased peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α) expression in skeletal muscle in response to creatine depletion.
Knowing these similarities, Winder and Hardie et al. designed an experiment to see if AMPK was influenced by thyroid hormone.
[55][56][57] They found that all of the subunits of AMPK were increased in skeletal muscle, especially in the soleus and red quadriceps, with thyroid hormone treatment.
[55] Loss of AMPK has been reported to alter the sensitivity of glucose sensing cells, through poorly defined mechanisms.
Loss of the AMPKα2 subunit in pancreatic β-cells and hypothalamic neurons decreases the sensitivity of these cells to changes in extracellular glucose concentration.
[58][59][60][61] Moreover, exposure of rats to recurrent bouts of insulin induced hypoglycemia/glucopenia, reduces the activation of AMPK within the hypothalamus, whilst also suppressing the counterregulatory response to hypoglycemia.
[65] A second AMPK-control system[66] localized to lysosomes depends on the Galectin-9-TAK1 system and ubiquitination responses at controlled by deubiquitinating enzymes such as USP9X leading to AMPK activation in response to lysosomal damage,[66] a condition that can occur biochemically, physically via protein aggregates such as proteopathic tau in Alzheimer's disease,[67][68] crystalline silica causing silicosis,[68] cholesterol crystals causing inflammation via NLRP3 inflammasome and rupture of atherosclerotic lesions,[69] urate crystals associated with gout, or during microbial invasion such as Mycobacterium tuberculosis[68][70] or coronaviruses causing SARS.
[71] Both of the above lysosomally localized systems controlling AMPK activate it in response to metformin,[66][72] a widely prescribed anti-diabetic drug.
[17] Additionally, studies where the AMPK activator metformin was used to treat diabetes found a correlation with a reduced risk of cancer, compared to other medications.
[73] In contrast, some studies have linked AMPK with a role as a tumor promoter by protecting cancer cells from stress.
[74] A seemingly paradoxical role of AMPK occurs when we take a closer look at the energy-sensing enzyme in relation to exercise and long-term training.