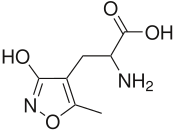
AMPA receptor
[12] S831 is phosphorylated by CaMKII and PKC during LTP, which helps deliver GluA1-containing AMPAR to the synapse,[13] and increases their single channel conductance.
The mechanism of desensitization is believed to be due to a small change in angle of one of the parts of the binding site, closing the pore.
[20] AMPARs open and close quickly (1ms), and are thus responsible for most of the fast excitatory synaptic transmission in the central nervous system.
Here, A→I editing alters the uncharged amino acid glutamine (Q) to the positively charged arginine (R) in the receptor's ion channel.
The positively charged amino acid at the critical point makes it energetically unfavourable for calcium to enter the cell through the pore.
[23] Alongside RNA editing, alternative splicing allows a range of functional AMPA receptor subunits beyond what is encoded in the genome.
[27] The flip form is present in prenatal AMPA receptors and gives a sustained current in response to glutamate activation.
In fact, it has been shown that, following a single paired-avoidance paradigm in mice, LTP can be recorded in some hippocampal synapses in vivo.
[29] The molecular basis for LTP has been extensively studied, and AMPARs have been shown to play an integral role in the process.
Ligand binding causes the AMPARs to open, and Na+ flows into the postsynaptic cell, resulting in a depolarization.
NMDARs can open only when a depolarization from the AMPAR activation leads to repulsion of the Mg2+ cation out into the extracellular space, allowing the pore to pass current.
The Ca2+ that enters the cell triggers the upregulation of AMPARs to the membrane, which results in a long-lasting increase in EPSP size underlying LTP.
The first key step in the process following glutamate binding to NMDARs is the influx of calcium through the NMDA receptors and the resultant activation of Ca2+/calmodulin-dependent protein kinase (CaMKII).
One possibility is that, during LTP, there is lateral movement of AMPA receptors from perisynaptic sites directly to the PSD.
Disruption of myosin Vb interaction with Rab11 and Rab11-FIP2 blocks spine growth and AMPA receptor trafficking.
[44] Therefore, it is possible that myosin may drive the lateral movement of AMPA receptors in the perisynaptic region to the PSD.
[11] The motion of AMPA receptors on the synaptic membrane are well approximated as a Brownian, which can however be stabilized at the PSD by retention forces.
[13] Under basal conditions, the regulated pathway is essentially inactive, being transiently activated only upon the induction of long-term potentiation.
[39][56] The GluA2 C terminus interacts with and binds to N-ethylmaleimide sensitive fusion protein,[57][58][59] which allows for the rapid insertion of GluR2-containing AMPA receptors at the synapse.
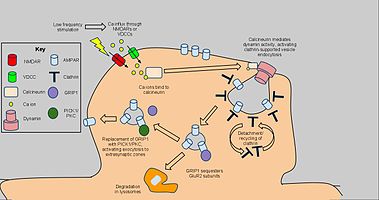
[61][62][63] Long-term depression enacts mechanisms to decrease AMPA receptor density in selected dendritic spines, dependent on clathrin and calcineurin and distinct from that of constitutive AMPAR trafficking.
The starting signal for AMPAR endocytosis is an NMDAR-dependent calcium influx from low-frequency stimulation, which in turn activates protein phosphatases PP1 and calcineurin.
[64] Blockage of PP1 did not prevent AMPAR endocytosis, but antagonist application to calcineurin led to significant inhibition of this process.
[65] Calcineurin interacts with an endocytotic complex at the postsynaptic zone, explaining its effects on LTD.[66] The complex, consisting of a clathrin-coated pit underneath a section of AMPAR-containing plasma membrane and interacting proteins, is the direct mechanism for reduction of AMPARs, in particular GluR2/GluR3 subunit-containing receptors, in the synapse.
[70] Nevertheless, the highlighted calcium-dependent, dynamin-mediated mechanism above has been implicated as a key component of LTD. and as such may have applications to further behavioral research.
[76] [77] Perampanel (trade name: Fycompa) received Marketing Authorisation Approval by the European Commission for the treatment of partial epilepsy on July 27, 2012.
[78] This direct inhibition of excitatory neurotransmission by decanoic acid in the brain contributes to the anticonvulsant effect of the medium-chain triglyceride ketogenic diet.
[78][79] Preclinical research suggest that several derivatives of aromatic amino acids with antiglutamatergic properties including AMPA receptor antagonism and inhibition of glutamate release such as 3,5-dibromo-D-tyrosine and 3,5-dibromo-L-phenylalnine exhibit strong anticonvulsant effect in animal models suggesting use of these compounds as a novel class of antiepileptic drugs.