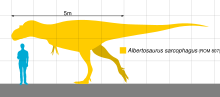
Albertosaurus
Albertosaurus (/ælˌbɜːrtəˈsɔːrəs/; meaning "Alberta lizard") is a genus of large tyrannosaurid theropod dinosaur that lived in northwestern North America during the early to middle Maastrichtian age of the Late Cretaceous period, about 71 million years ago.
[2][3] As a tyrannosaurid, Albertosaurus was a bipedal predator with short arms, two-fingered hands, and a massive head with dozens of large, sharp teeth, a strong sense of smell, powerful binocular vision, and a bone crushing bite force.
[6][7] The type specimen is a partial skull collected on June 9, 1884, from an outcrop of the Horseshoe Canyon Formation alongside the Red Deer River in Alberta.
In 1889, Tyrrell's colleague Thomas Chesmer Weston found an incomplete smaller skull associated with some skeletal material at a location nearby.
[12] Shortly later, Osborn pointed out that D. incrassatus was based on generic tyrannosaurid teeth, so the two Horseshoe Canyon skulls could not be confidently referred to that species.
The Horseshoe Canyon skulls also differed markedly from the remains of D. aquilunguis, type species of Dryptosaurus, so Osborn gave them the new name Albertosaurus sarcophagus in 1905.
By the early twenty-first century, some concerns had arisen that, due to the damaged state of the holotype, Albertosaurus might be a nomen dubium that could only be used for the type specimen itself because other fossils could not reliably be assigned to it.
However, in 2010, Thomas Carr established that the holotype, the paratype, and comparable later finds all shared a single common unique trait, or autapomorphy.
[13] On August 11, 1910, American paleontologist Barnum Brown discovered the remains of a large group of Albertosaurus at another quarry alongside the Red Deer River.
Palaeontologist Philip J. Currie of the Royal Tyrrell Museum of Palaeontology rediscovered the bonebed in 1997 and resumed fieldwork at the site, which is now located inside Dry Island Buffalo Jump Provincial Park.
[16] In 1911, Barnum Brown, during the second year of the American Museum of Natural History's operations in Alberta, uncovered a fragmentary partial Albertosaurus skull at the Red Deer River near Tolman Bridge (specimen AMNH 5222).
[19] In 2003, Philip J. Currie, benefiting from much more extensive finds and a general increase in anatomical knowledge of theropods, compared several tyrannosaurid skulls and came to the conclusion that the two species are more distinct than previously thought.
In 1988, Gregory S. Paul based Albertosaurus megagracilis on a small tyrannosaurid skeleton, specimen LACM 28345, from the Hell Creek Formation of Montana.
[28] Wide openings in the skull, called fenestrae, provided space for muscle attachment sites and sensory organs that reduced its overall weight.
[20] Like with Tyrannosaurus rex, the maxillary (cheek) teeth of Albertosaurus were adapted in general form to resist lateral forces exerted by a struggling prey animal.
[39] Unlike tyrannosaurs, more ancient predators, like phytosaurs and Dimetrodon, had no adaptations to prevent the crack-like serrations of their teeth from spreading when subjected to the forces of feeding.
Compared with the more robust tyrannosaurines, albertosaurines had slender builds, with proportionately smaller skulls and longer bones of the lower legs (tibia) and feet (metatarsals and phalanges).
Using bone histology, the age of an individual animal at the time of death can often be determined, allowing growth rates to be estimated and compared with other species.
The youngest known Albertosaurus is a two-year-old discovered in the Dry Island bonebed, which would have weighed about 50 kilograms (110 lb) and measured slightly more than 2 metres (6.6 feet) long.
[22][44] During growth, thickening of the tooth morphology changed so much that, had the association of young and adult skeletons on the Dry Island bonebed not proven that they belonged to the same taxon, the teeth of juveniles would likely have been identified by statistical analysis as those of a different species.
[6] A hypothesis of Albertosaurus life history postulates that hatchlings died in large numbers, but have not been preserved in the fossil record because of their small size and fragile construction.
This elevated mortality rate continued throughout adulthood, perhaps due to the high physiological demands of procreation, including stress and injuries received during intraspecific competition for mates and resources, and the eventual, ever-increasing effects of senescence.
The comparison with modern animals and other tyrannosaurids lends support to this life history hypothesis, but bias in the fossil record may still play a large role, especially since more than two-thirds of all Albertosaurus specimens are known from the exact same locality.
[14] Others have speculated that, instead of social groups, at least some of these finds represent Komodo dragon-like mobbing of carcasses, where aggressive competition leads to some of the predators being killed and even cannibalized.
[20] A similar situation is observed in modern Komodo dragons, with hatchlings beginning life as small insectivores before growing to become the dominant predators on their islands.
In 2010, Currie, though still favouring the hunting pack hypothesis, admitted that the concentration could have been brought about by other causes, such as a slowly rising water level during an extended flood.
[56] In 2009, researchers hypothesized that smooth-edged holes found in the fossil jaws of tyrannosaurid dinosaurs, such as Albertosaurus, were caused by a parasite similar to Trichomonas gallinae, which infects birds.
These younger units of this geologic formation date to the early Maastrichtian age of the Late Cretaceous period, about 70 to 68 million years ago.
Like most of the other vertebrate fossils from the formation, Albertosaurus remains are found in deposits laid down in the deltas and floodplains of large rivers during the later half of Horseshoe Canyon times.
The saltwater plesiosaur Leurospondylus has been found in marine sediments in the Horseshoe Canyon, while freshwater environments were populated by turtles, Champsosaurus, and crocodilians like Leidyosuchus and Stangerochampsa.