Arabinogalactan protein
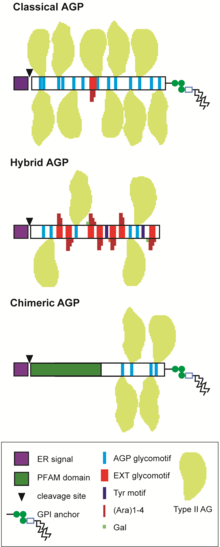
[3][4] AGPs belong to large multigene families and are divided into several sub-groups depending on the predicted protein sequence.
Some AGPs are rich in uronic acids (GlcA), resulting in a charged polysaccharide moiety, and others have short oligosaccharides of Araf.
[23] Specific sets of hydroxyproline O-β-galactosyltransferases, β-1,3-galactosyltransferases, β-1,6-galactosyltransferases, α-arabinosyltransferases, β-glucuronosyltransferases, α-rhamnosyltransferases, and α- fucosyltransferases are responsible for the synthesis of these complex structures.
Seven residues and longer are sufficient for cross-linking, leading to precipitation of the glycans with the Yariv phenylglycosides, which are observed with classical AGPs binding to β-Yariv dyes.
[26] After translation, the AGP protein backbones are highly decorated with complex carbohydrates, primarily type II AG polysaccharides.
[27] The biosynthesis of the mature AGP involves cleavage of the signal peptide at the N-terminus, hydroxylation on the P residues and subsequent glycosylation and in many cases addition of a GPI-anchor.
These include the assembly of tri-mannose (Man), galactose, non-N-acetylated glucosamine (GlcN) and ethanolamine phosphate to form the mature GPI moiety.
[41][43][44][40][10][45] The only plant GPI anchor structure characterized to date is the GPI-anchored AGP from Pyrus communis suspension-cultured cells.
[46] As proposed by Oxley and Bacic,[40] the GPI-anchored AGPs are likely released via cleavage by some phospholipases (PLs) (C or D) and secreted into the extracellular compartment.
Bioinformatics analysis using mammalian β-1,3-galactosyltransferase (GalT) sequences as templates suggested involvement of the Carbohydrate-Active enZYmes (CAZy) glycosyltransferase (GT) 31 family in the synthesis of the galactan chains of the AG backbone.
[47] A GT31 Clade 10 gene, KNS4/UPEX1, encodes a β-1,3-GalT capable of synthesizing β-1,3-Gal linkages found in type II AGs present in AGPs and/or pectic rhamnogalacturonan I (RG-I).
[51] Another GT31 Clade 10 member, named GALT31A, encodes a β-1,6-GalT when heterologously expressed in E. coli and Nicotiana benthamiana and elongated β-1,6-galactan side chains of AGP glycans.
[53] Three members of GT14 named GlcAT14A, GlcAT14B, and GlcAT14C were reported to add GlcA to both β-1,6- and β-1,3-Gal chains in an in vitro enzyme assay following heterologous expression in Pichia pastoris.
[55] A GT77 family member, REDUCED ARABINOSE YARIV (RAY1), was found to be a β-arabinosyltransferase that adds a β-Araf to methyl β-Gal of a Yariv-precipitable wall polymer.
[57] More research is expected to functionally identify other genes involved in AGP glycosylation and their interactions with other plant cell wall components.
Human uses of AGPs include the use of gum arabic in the food and pharmaceutical industries because of natural properties in thickening and emulsification.
[58][59] AGPs in cereal grains have potential applications in biofortification,[60] as sources of dietary fibre to support gut bacteria[61] and protective agents against ethanol toxicity.
[66][67][68] Conventional methods to study functions of AGPs include the use of β-glycosyl (usually glucosyl) Yariv reagents and monoclonal antibodies (mAbs).
[80][81][82] Another possible mechanism, largely based on the study of FLAs, suggests the combination of fasciclin domain and AG glycans can mediate cell-cell adhesion.
[83][84] The functions of AGPs in plant growth and development processes rely heavily on the incredible diversity of their glycan and protein backbone moieties.
Human uses of AGPs include the use of Gum arabic in the food and pharmaceutical industries because of natural properties in thickening and emulsification.
[58][59] AGPs in cereal grains have potential applications in biofortification,[60] as sources of dietary fibre to support gut bacteria[61] and protective agents against ethanol toxicity.
[62] This article was adapted from the following source under a CC BY 4.0 license (2021) (reviewer reports): Yingxuan Ma; Kim Johnson (15 January 2021).