[7] Molecular recognition takes place in a noisy, crowded biological environment and the recognizer often has to cope with the task of selecting its target among a variety of similar competitors.
The group of C. Dekker (Delft University) directly probed the interactions involved in homology search by combining magnetic and optical tweezers.
The data indicate a physical picture for homology recognition in which the fidelity of the search process is governed by the distance between the DNA-binding sites.
The authors conclude that their interpretation of the measurements "is akin to a conformational proofreading scheme ... where the dsDNA, and not the RecA filament, is the active, recognizing search entity.
The ribosome is a complex molecular machine that, in order to synthesize proteins during the translation process, has to decode mRNAs by pairing their codons with matching tRNAs.
Decoding is a major determinant of fitness and requires accurate and fast selection of correct tRNAs among many similar competitors.
[11] The research focused on the question of how DNA-repair proteins scan the human genome for UV-induced damage during the initial step of nucleotide excision repair (NER).
Analysis of the rates of dissociation for the transient binding molecules on both undamaged and damaged DNA show multiple dwell times over three orders of magnitude...
The authors conclude from their detailed kinetic measurements that UV-DDB recognizes lesions using a conformational proofreading mechanism via multiple intermediates.
In the kinetic proofreading[12][13] schema, a time delay (equivalently, an irreversible intermediate stage) is introduced during the formation of the correct or incorrect complexes.
Conformational proofreading in homologous recombination
.
Top
: The binding probability to homologous (correct) and non-homologous (wrong) DNA sequences decrease with the deformation
energy barrier
. The wrong binding probability decreases before the correct one.
Bottom
: As a result, the
Fitness
which is the difference, Fitness = Prob(Correct) − Prob(Wrong), is maximal at a non-zero deformation energy. This barrier is optimal in the sense that it significantly reduces the binding probability while keeping the correct binding probability roughly the same. Biochemical measurements of RecA-induced recombination suggest that the observed deformation is nearly optimal.
[
3
]
[
4
]
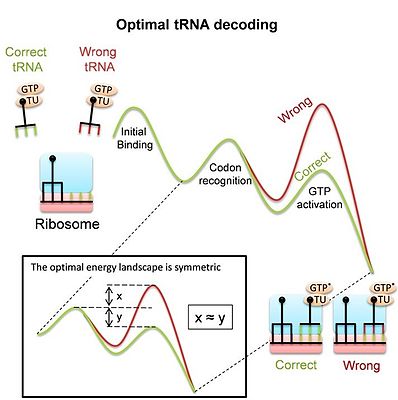
Ribosome uses conformational proofreading for tRNA decoding
.
main
: The curves show the free energy landscape of codon recognition as suggested by experiments. In the stages that are sensitive to codon identity, the pathways of correct (green) and wrong (red) tRNAs split. The multistage kinetics include: Initial binding and codon recognition: a complex of elongation factor (EF-Tu) and aminoacyl-tRNA binds to the ribosome. Codon is recognized by pairing with the anticodon, and by additional interaction with the "decoding center" of the ribosome. As a result, correct (cognate) tRNAs are more stable than non-cognate ones. GTP activation and hydrolysis: Codon recognition leads to global conformational changes of the ribosome and tRNA, which are different for cognate or non-cognate tRNAs and affect GTP activation and hydrolysis by EF-Tu.
The conformational proofreading model explains these conformational changes as a means to enhance tRNA recognition.
inset
: The
symmetric
adapted landscape implies that the ratio of forward and backward rates is inverted between the correct and wrong energy landscapes.
[
5
]