Dioxygenase
Aerobic life, from simple single-celled bacteria species to complex eukaryotic organisms, has evolved to depend on the oxidizing power of dioxygen in various metabolic pathways.
From energetic adenosine triphosphate (ATP) generation to xenobiotic degradation, the use of dioxygen as a biological oxidant is widespread and varied in the exact mechanism of its use.
The majority of dioxygenases fully incorporate dioxygen into a single substrate, and a variety of cofactor schemes are utilized to achieve this.
For example, in the α-ketoglutarate-dependent enzymes, one atom of dioxygen is incorporated into two substrates, with one always being α-ketoglutarate, and this reaction is brought about by a mononuclear iron center.
The most widely observed cofactor involved in dioxygenation reactions is iron, but the catalytic scheme employed by these iron-containing enzymes is highly diverse.
Despite this common oxygenation event, the mononuclear iron dioxygenases are diverse in how dioxygen activation is used to promote certain chemical reactions.
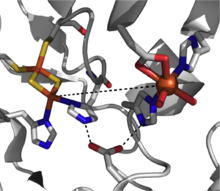
[1][2][3] Most mononuclear iron dioxygenases are members of the cupin superfamily in which the overall domain structure is described as a six-stranded β-barrel fold (or jelly roll motif).
[2][4] Extradiol members utilize ferrous iron as the active redox state, and this center is commonly coordinated octahedrally through a 2-His-1-Glu motif with labile water ligands occupying empty positions.
[2][4][7] This activated oxygen species then proceeds to react with the substrate ultimately cleaving the carbon-carbon bond adjacent to the hydroxyl groups through the formation of an α-keto lactone intermediate.
[6] Upon oxygen binding, a poorly understood transformation occurs during which 2OG is oxidatively decarboxylated to succinate and the O-O bond is cleaved to form a Fe(IV)-oxo (ferryl) intermediate.
This radical then couples to the hydroxide ligand, producing the hydroxylated product and the Fe(II) resting state of the enzyme.
[2] These enzymes are prominently found in soil bacteria such as Pseudomonas,[3] and their reactions constitute the initial step in the biodegradation of aromatic hydrocarbons.
In this respect, these enzymes are reminiscent of the intradiol catechol dioxygenases whereby the metal centers activate the substrate for subsequent reaction with dioxygen.
[16] These enzymes catalyze the degradation of quinolone heterocycles in a manner similar to quercetin dioxygenase, but are thought to mediate a radical reaction of a dioxygen molecule with a carbanion on the substrate (figure 5).