ESCRT
[1][2] These ESCRT components have been isolated and studied in a number of organisms including yeast and humans.
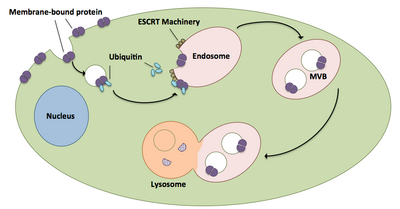
Multivesicular body (MVB) biogenesis is a process in which ubiquitin-tagged proteins enter organelles called endosomes via the formation of vesicles.
For example, abnormalities in ESCRT-III components can lead to neurological disorders such as hereditary spastic paraplegia (HSP).
[6] Cellular abscission, the process by which the membrane connecting two daughter cells is cleaved, is also mediated by ESCRT machinery.
Lastly, viral budding, or the process by which specific types of viruses exit cells, may not occur in the absence of ESCRT machinery.
Each of the ESCRT complexes and accessory proteins have unique structures that enable distinct biochemical functions.
[7] Once properly localized, these proteins are then taken into the endosome via vesicles, forming multivesicular bodies, and are eventually delivered to the lysosome where they are degraded.
This process is essential as it is the major pathway for the degradation of damaged proteins that have passed through the Golgi.
[3][6] Just after this ubiquitin E2 variant domain, a proline rich motif (GPPX3Y) is present that directs ESCRT-I to the midbody during membrane abscission.
[1] The ESCRT-II complex functions primarily during the biogenesis of multivesicular bodies and delivery of ubiquitin tagged proteins to the endosome.
[15] These filamentous structures are also present during multivesicular body formation and function as a ring-like fence that plugs the budding vesicle to prevent cargo proteins from escaping into the cell's cytosol.
[3] The Vps4-Vta1 proteins are required for the stripping of other ESCRT components (usually ESCRT-III) from membranes once a particular process has been completed.
Though not essential, Vta1 has been shown to aid in Vps4 ring assembly, accelerate the ATPase activity of Vsp4, and encourage ESCRT-III disassembly.
[19] This results in the removal of ubiquitin tags from proteins targeted for degradation in the lysosome just prior to the generation of multivesicular bodies.
[20] Multivesicular bodies play a large role in the transport of ubiquitinated proteins and receptors to a lysosome.
[21] ESCRT complexes transport ubiquitinated cargo to cellular vesicles that bud directly into the cell’s endosomal compartment, forming multivesicular bodies.
Since it is conserved in a number of archaea, membrane abscission is considered to be the earliest role for ESCRT machinery.
[6][23] Cep55 then recruits the Vps23 subunit of ESCRT-I and accessory protein ALIX, which form into rings on either side of the midbody.
[1][12][19] The formation of this spiral-like structure deforms the membrane and the AAA-ATPase spastin is brought in by Did2 and Ist1 to cleave the microtubules formed at the midbody.