Homologous chromosome
[2] Chromosomes are linear arrangements of condensed deoxyribonucleic acid (DNA) and histone proteins, which form a complex called chromatin.
[3] Early in the 1900s, William Bateson and Reginald Punnett were studying genetic inheritance and they noted that some combinations of alleles appeared more frequently than others.
Using test cross experiments, he revealed that, for a single parent, the alleles of genes near to one another along the length of the chromosome move together.
[2] Creighton and McClintock discovered that the new allele combinations present in the offspring and the event of crossing over were directly related.
[5] The actual length of the arm, in accordance with the gene locations, is critically important for proper alignment.
Centromere placement on the chromosome can be characterized by four main arrangements, either metacentric, submetacentric, acrocentric, or telocentric.
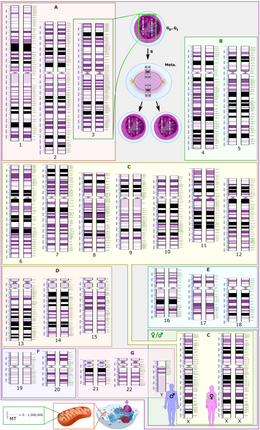
In humans, the 22 pairs of homologous autosomal chromosomes contain the same genes but code for different traits in their allelic forms, as one was inherited from the mother and one from the father.
[7] During meiosis, genetic recombination (by random segregation) and crossing over produces daughter cells that each contain different combinations of maternally and paternally coded genes.
In prophase I, the DNA has already undergone replication so each chromosome consists of two identical chromatids connected by a common centromere.
[10] This pairing occurs by a synapsis process where the synaptonemal complex – a protein scaffold – is assembled and joins the homologous chromosomes along their lengths.
[10] In addition, another type of recombination referred to as synthesis-dependent strand annealing (SDSA) frequently occurs.
In the process of crossing-over, genes are exchanged by the breaking and union of homologous portions of the chromosomes' lengths.
[7] Both the non-crossover and crossover types of recombination function as processes for repairing DNA damage, particularly double-strand breaks.
At the diplotene stage of prophase I the synaptonemal complex disassembles before which will allow the homologous chromosomes to separate, while the sister chromatids stay associated by their centromeres.
[10] Instead, the replicants, or sister chromatids, will line up along the metaphase plate and then separate in the same way as meiosis II – by being pulled apart at their centromeres by nuclear mitotic spindles.
Faulty segregation can lead to fertility problems, embryo death, birth defects, and cancer.
If this uneven division occurs in meiosis I, then none of the daughter cells will have proper chromosomal distribution and non-typical effects can ensue, including Down's syndrome.
[5] While the main function of homologous chromosomes is their use in nuclear division, they are also used in repairing double-strand breaks of DNA.
[16] Once the base pairs have been matched and oriented correctly between the two strands, the homologous chromosomes perform a process that is very similar to recombination, or crossing over as seen in meiosis.
[16] Current and future research on the subject of homologous chromosome is heavily focused on the roles of various proteins during recombination or during DNA repair.
Manipulating the repair function of homologous chromosomes might allow for bettering a cell's damage response system.