Hydrophobic mismatch
In order to avoid unfavorable exposure of hydrophobic surfaces to a hydrophilic environment, biological membrane tends to adapt to such mismatch.
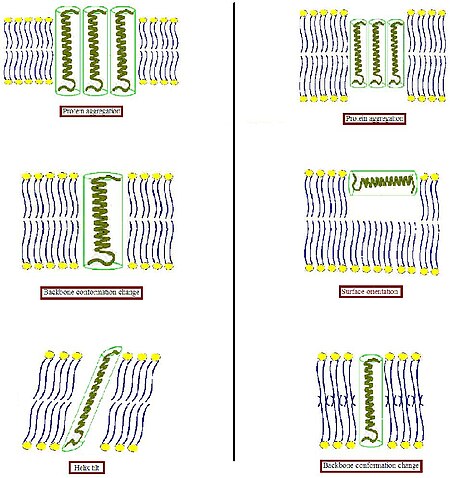
[10][11][12] Electron microscopy studies on bacteriorhodopsin, reconstituted in saturated and unsaturated fluid PC bilayers with varying chain length, showed that protein aggregation occurred only with a rather large mismatch, and that bilayer thicknesses of 4 angstrom thicker and 10 angstrom thinner than the estimated hydrophobic thickness of the protein are allowed without induction of significant aggregation.
In large proteins that span the membrane multiple times, changes in helical tilt may occur with little effect on lipid packing.
The experimental evidence was shown by a fluorescence study on an artificial peptide with a 19 amino acid long hydrophobic sequence of mainly leucines and flanked on both sides with lysines as anchoring residues.
[15] The results indicated that a conversion from a dominant transmembrane to parallel orientation of the peptide could be induced by modulating bilayer thickness via addition of cholesterol or by increasing lipid chain length.
In their model, they relate the energy stored in the undulations of the membrane surface caused by the mismatch to the elastic properties of the lipids and proteins.
The interaction potentials are estimated based on experimental data derived from thermodynamic and mechanical measurements of membrane properties.
[21] A molecular modeling was combined with phenomenological free energy contributions describing lipid head group repulsion and membrane solvent surface tension.
In recent years, great advances in X-ray crystallography and electron microscopy techniques have yielded new insights of the lipid interactions of larger proteins.
[28][29] Rafts have putative roles in many physiological processes, such as signal transduction, endocytosis, apoptosis, protein trafficking, and lipid regulation.