Karyogamy
Karyogamy is the final step in the process of fusing together two haploid eukaryotic cells, and refers specifically to the fusion of the two nuclei.
[1] Haploid organisms such as fungi, yeast, and algae can have complex cell cycles, in which the choice between sexual or asexual reproduction is fluid, and often influenced by the environment.
Some organisms, in addition to their usual haploid state, can also exist as diploid for a short time, allowing genetic recombination to occur.
In haploid organisms that lack sexual cycles, karyogamy can also be an important source of genetic variation during the process of forming somatic diploid cells.
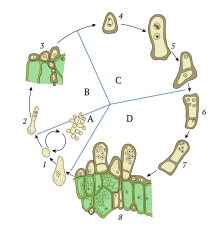
[2] The role of karyogamy in sexual reproduction can be demonstrated most simply by single-celled haploid organisms such as the algae of genus Chlamydomonas or the yeast Saccharomyces cerevisiae.
[3] When subjected to environmental stress, such as nitrogen starvation in the case of Chlamydomonas, cells are induced to form gametes.
[4] Gamete formation in single-celled haploid organisms such as yeast is called sporulation, resulting in many cellular changes that increase resistance to stress.
Gamete formation in multicellular fungi occurs in the gametangia, an organ specialized for such a process, usually by meiosis.
[7][8] Mate recognition in the simplest eukaryotes is achieved through pheromone signaling, which induces shmoo formation (a projection of the cell) and begins the process of microtubule organization and migration.
Pheromones used in mating type recognition are often peptides, but sometimes trisporic acid or other molecules, recognized by cellular receptors on the opposite cell.
One possible advantage of sexual reproduction is that it results in more genetic variability, providing the opportunity for adaptation through natural selection.
Thus, karyogamy is the key step in bringing together a variety of genetic material in order to ensure recombination in meiosis.
In the sliding cross-bridge model, the microtubules run antiparallel to each other for the entire distance between the two pronuclei, forming cross-links to each other, and each attaching to the opposite nucleus at the plus end.
In either model, it is believed that microtubule shortening occurs at the plus end and requires Kar3p (in yeast), a member of a family of kinesin-like proteins.
[13] As described above, the reorganization of accessory and motor proteins during pronuclear migration also serves to orient the spindle pole bodies in the correct direction for efficient nuclear congression.
The second mechanism, involving karyogamy of somatic cells, can produce heterozygous diploids if the two nuclei differ in genetic information.
The formation of somatic diploids is generally rare, and is thought to occur because of a mutation in the karyogamy repressor gene (KR).