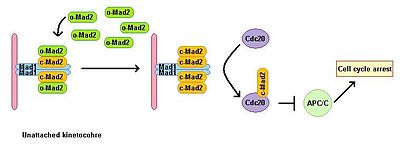
Mad1
They are conserved in all eukaryotes and are involved in a pathway that is active in prometaphase to prevent the premature separation of sister chromatids and constitute the so-called spindle assembly checkpoint (SAC).
This checkpoint monitors the status of chromosome attachment to the mitotic spindle and inhibits the metaphase to anaphase transition by preventing the activation of the anaphase-promoting complex/cyclosome (APC/C), and thereby the degradation of cell cycle regulators.
Mad1 recruits the important SAC component Mad2 to unattached kinetochores and induces mitotic arrest signal amplification.
[3] If a kinetochore is attached to spindle, SAC inhibitor p31comet inhibits Mad1 mediated conformational rearrangement of Mad2 and prevents Mad2 from binding to Cdc20.
Elongated Mad1 monomers are tightly held together by a parallel coiled-coil involving the N-terminal alpha helices.
The first part of this binding domain is flexible and adopts different conformations giving rise to an asymmetric complex.
In their work, employing thermodynamic studies, Sironi et al.[2] show that Mad1 functions such as to slow down the rate of Mad2-Cdc20 complex formation and therefore acts as a competitive inhibitor in vivo.
The authors therefore conclude that an unperturbed Mad1-Mad2 complex will not release Mad2 requiring a novel, so far poorly understood, mechanism of conformational change.
[2] Mismatches in chromosome number (aneuploidies) during meiosis are responsible for human diseases like Down's syndrome and also emerge frequently in cancer cells.