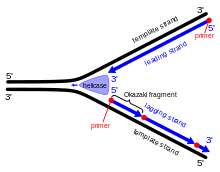
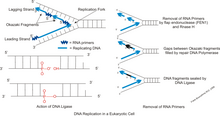
Okazaki fragments
The primase and polymerase move in the opposite direction of the fork, so the enzymes must repeatedly stop and start again while the DNA helicase breaks the strands apart.
[4] [2] During the 1960s, Reiji and Tsuneko Okazaki conducted experiments involving DNA replication in the bacterium Escherichia coli.
3' and 5' are specifically numbered carbons on the deoxyribose ring in nucleic acids, and refer to the orientation or directionality of a strand.
The hypothesis was further supported by the discovery of polynucleotide ligase, an enzyme that links short DNA strands together.
[6] In 1968, Reiji and Tsuneko Okazaki gathered additional evidence of nascent DNA strands.
The cells infected with the T4 phages accumulated a large number of short, newly synthesized DNA chains, as predicted in the hypothesis, when exposed to high temperatures.
Pol δ frequently encounters the downstream primed Okazaki fragment and displaces the RNA/DNA initiator primer into a 5′ flap.
show that in the FEN1 suggest a ‘tracking; model where the nuclease moves from the 5’ flap to its base to preform cleavage.
The cleavage is inhibited when the 5’ end of the DNA flap is blocked either with a complementary primer or a biotin-conjugated streptavidin moiety.
Sequential strand displacement and cleavage by Pol δ and FEN1, respectively, helps to remove the entire initiator RNA before ligation.
[9] Primase adds RNA primers onto the lagging strand, which allows synthesis of Okazaki fragments from 5' to 3'.
DNA polymerase on the lagging strand also has to be continually recycled to construct Okazaki fragments following RNA primers.
Last function of DNA polymerase δ is to serve as a supplement to FEN1/RAD27 5’ Flap Endonuclease activity.
The RAD27/FEN1 creates nicks between adjacent Okazaki fragments by minimizing the amount of strand-expulsion in the lagging strand.
Further research implicates the supplementary role of proliferating cell nuclear antigen (PCNA) to DNA ligase I's function of joining Okazaki fragments.
Then, DNA ligase I binds to the PCNA, which is clamped to the nicks of the lagging strand, and catalyzes the formation of phosphodiester bonds.
Thus, FEN1's function is necessary to Okazaki fragment maturation in forming a long continuous DNA strand.
Likewise, during DNA base repair, the damaged nucleotide is displaced into a flap and subsequently removed by FEN1.
[15][16] Dna2 endonuclease does not have a specific structure and their properties are not well characterized, but could be referred as single-stranded DNA with free ends (ssDNA).
Also, Dna2 endonuclease has a pivotal role in the intermediates created during diverse DNA metabolisms and is functional in telomere maintenance.
In the presence of a single stranded DNA-binding protein RPA, the DNA 5' flaps become too long, and the nicks no longer fit as substrate for FEN1.
Since this process is so common, Okazaki maturation will take place around a million times during one completion of DNA replication.
This means that the piecewise generation of Okazaki fragments can keep up with the continuous synthesis of DNA on the leading strand.
We take from this that prokaryotic cells are simpler in structure, they have no nucleus, organelles, and very little of DNA, in the form of a single chromosome.
For example, a mutation related to primase affects RNA primer removal and can make the DNA strand more fragile and susceptible to breaks.
Another mutation concerns polymerase α, which impairs the editing of the Okazaki fragment sequence and incorporation of the protein into the genetic material.
Both alterations can lead to chromosomal aberrations, unintentional genetic rearrangement, and a variety of cancers later in life.
The homozygous knockout mutant mice experienced a "failure of cell proliferation" and "early embryonic lethality" (27).
This is because the FFAA mutation prevents the FEN1 from interacting with PCNA (proliferating cell nuclear antigen), consequently not allowing it to complete its purpose during Okazaki fragment maturation.
In time, these nicks also cause full chromosome breaks, which could lead to severe mutations and cancers.