Ribonuclease H
Ribonuclease H (abbreviated RNase H or RNH) is a family of non-sequence-specific endonuclease enzymes that catalyze the cleavage of RNA in an RNA/DNA substrate via a hydrolytic mechanism.
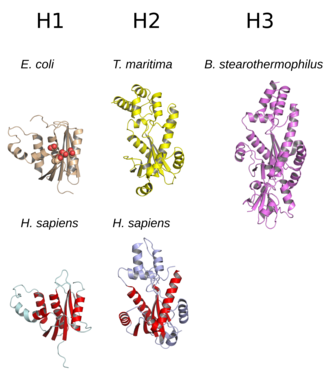
The family is divided into evolutionarily related groups with slightly different substrate preferences, broadly designated ribonuclease H1 and H2.
Human ribonuclease H2 is a heterotrimeric complex composed of three subunits, mutations in any of which are among the genetic causes of a rare disease known as Aicardi–Goutières syndrome.
[4] Additionally, RNase H1-like retroviral ribonuclease H domains occur in multidomain reverse transcriptase proteins, which are encoded by retroviruses such as HIV and are required for viral replication.
[2][7] Prokaryotic HIII enzymes are members of the broader H2 group and share most structural features with H2, with the addition of an N-terminal TATA box binding domain.
[21][22] Ribonuclease H1 enzymes require at least four ribonucleotide-containing base pairs in a substrate and cannot remove a single ribonucleotide from a strand that is otherwise composed of deoxyribonucleotides.
For this reason, it is considered unlikely that RNase H1 enzymes are involved in the processing of RNA primers from Okazaki fragments during DNA replication.
In knockout mouse models, RNase H1-null mutants are lethal during embryogenesis due to defects in replicating mitochondrial DNA.
[29][30][28] Although both H1 and H2 are present in the mammalian cell nucleus, H2 is the dominant source of RNase H activity there and is important for maintaining genome stability.
[32] The active site of nearly all RNases H contains four negatively charged amino acid residues, known as the DEDD motif; often a histidine e.g. in HIV-1, human or E. coli is also present.
[11][33][34] Based on experimental evidence and computer simulations the enzyme activates a water molecule bound to one of the metal ions with the conserved histidine.
[33][35] The transition state is associative in nature [17] and forms an intermediate with protonated phosphate and deprotonated alkoxide leaving group.
Experimental evidence from time-resolved crystallography and similar nucleases points to a role of a third ion in the reaction recruited to the active site.
[39][40][41] In small studies, mutations in human RNase H1 have been associated with chronic progressive external ophthalmoplegia, a common feature of mitochondrial disease.
[25] Mutations in any of the three RNase H2 subunits are well-established as causes of a rare genetic disorder known as Aicardi–Goutières syndrome (AGS),[3] which manifests as neurological and dermatological symptoms at an early age.
[43] The symptoms of AGS closely resemble those of congenital viral infection and are associated with inappropriate upregulation of type I interferon.
[45] Mutations in 2B have been associated with somewhat milder neurological impairment[46] and with an absence of interferon-induced gene upregulation that can be detected in patients with other AGS-associated genotypes.
Inhibitors of retroviral RNase H of several different chemotypes have been identified, many of which have a mechanism of action based on chelation of the active-site cations.
[7][32] RNase HIII, which is unique to prokaryotes, has a scattered taxonomic distribution and is found in both bacteria and archaea;[32] it is believed to have diverged from HII fairly early.
[60][61] RNase HII can be used to degrade the RNA primer component of an Okazaki fragment or to introduce single-stranded nicks at positions containing a ribonucleotide.
[26][63][64] RNase H activity was subsequently discovered in E. coli[65] and in a sample of oncoviruses with RNA genomes during early studies of viral reverse transcription.
[74] However, the yeast B and C subunits have very low sequence identity to their homologs in other organisms, and the corresponding human proteins were conclusively identified only after mutations in all three were found to cause Aicardi–Goutières syndrome.