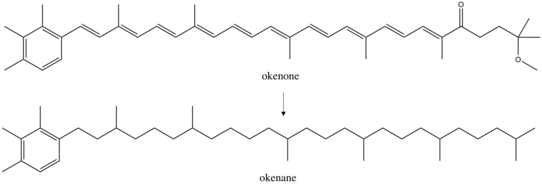
Okenane
Discovery of okenane in marine sediments implies a past euxinic environment, where water columns were anoxic and sulfidic.
This is potentially tremendously important for reconstructing past oceanic conditions, but so far okenane has only been identified in one Paleoproterozoic (1.6 billion years old) rock sample from Northern Australia.
Okenone appears to be best adapted to the yellow-green transition (520 nm) of the visible spectrum, capturing light below marine plankton in the ocean.
Finding any of these carotenoids in ancient rocks could constrain the depth of the oxic to anoxic transition as well as confine past ecology.
Okenane and chlorobactane discovered in Australian Paleoproterozoic samples allowed conclusions of a temporarily shallow anoxic transition, likely between 12 and 25m.
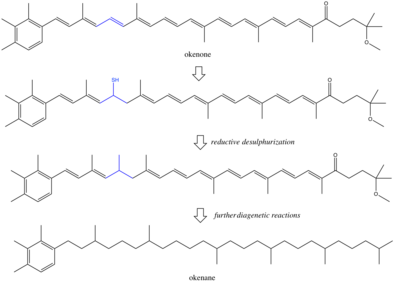
The exact reactions during diagenesis are poorly understood, although some have proposed reductive desulphurization as a mechanism for saturation of okenone to okenane.
[13] If Chromatiaceae gained the ability to create okenone more recently that the Paleoproterozoic, then the okenane does not track purple sulfur bacteria, but rather the original gene donor.
In modern environments, purple sulfur bacteria thrive in meromictic (permanently stratified) lakes[15] and silled fjords and are seen in few marine ecosystems.
[16] However, billions of years ago, when the oceans were anoxic and sulfidic, phototrophic sulfur bacteria had more habitable space.
Researchers at the Australian National University and the Massachusetts Institute of Technology investigated 1.6-billion-year-old rocks to examine the chemical conditions of the Paleoproterozoic ocean.
Many believe that this time had deeply penetrating oxic water columns because of the disappearance of banded iron formations roughly 1.8 billion years ago.
Examining rocks from the time uncovered biomarkers of both purple and green sulfur bacteria, adding evidence to support the Canfield Ocean hypothesis.