Cerebral cortex
It is the largest site of neural integration in the central nervous system,[2] and plays a key role in attention, perception, awareness, thought, memory, language, and consciousness.
[11] The cerebral cortex is folded in a way that allows a large surface area of neural tissue to fit within the confines of the neurocranium.
When unfolded in the human, each hemispheric cortex has a total surface area of about 0.12 square metres (1.3 sq ft).
For species of mammals, larger brains (in absolute terms, not just in relation to body size) tend to have thicker cortices.
[20] A genetic disorder of the cerebral cortex, whereby decreased folding in certain areas results in a microgyrus, where there are four layers instead of six, is in some instances seen to be related to dyslexia.
[22] Layer I consists largely of extensions of apical dendritic tufts of pyramidal neurons and horizontally oriented axons, as well as glial cells.
Inputs to the apical tufts are thought to be crucial for the feedback interactions in the cerebral cortex involved in associative learning and attention.
In the primary motor cortex of the frontal lobe, layer V contains giant pyramidal cells called Betz cells, whose axons travel through the internal capsule, the brain stem, and the spinal cord forming the corticospinal tract, which is the main pathway for voluntary motor control.
[34] In 1957, Vernon Mountcastle showed that the functional properties of the cortex change abruptly between laterally adjacent points; however, they are continuous in the direction perpendicular to the surface.
Cerebral veins drain the deoxygenated blood, and metabolic wastes including carbon dioxide, back to the heart.
The prenatal development of the cerebral cortex is a complex and finely tuned process called corticogenesis, influenced by the interplay between genes and the environment.
The most anterior (front, or cranial) part of the neural plate, the prosencephalon, which is evident before neurulation begins, gives rise to the cerebral hemispheres and later cortex.
In addition, laminar neurons are stopped in S or G2 phase in order to give a fine distinction between the different cortical layers.
[43] Although the majority of the cells that compose the cortex are derived locally from radial glia there is a subset population of neurons that migrate from other regions.
These GABAergic neurons are generated by progenitor cells in the medial ganglionic eminence (MGE) that migrate tangentially to the cortex via the subventricular zone.
During the first year of life the dendrites become dramatically increased in number, such that they can accommodate up to a hundred thousand synaptic connections with other neurons.
As the G1 phase of mitosis is elongated, in what is seen as selective cell-cycle lengthening, the newly born neurons migrate to more superficial layers of the cortex.
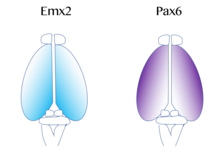
[53] The map of functional cortical areas, which include primary motor and visual cortex, originates from a 'protomap',[54] which is regulated by molecular signals such as fibroblast growth factor FGF8 early in embryonic development.
On the contrary, if mutations in Emx2 occur, it can cause the Pax6-expressing domain to expand and result in the frontal and motor cortical regions enlarging.
Therefore, researchers believe that similar gradients and signaling centers next to the cortex could contribute to the regional expression of these transcription factors.
Therefore, Fgf8 and other FGFs play a role in the regulation of expression of Emx2 and Pax6 and represent how the cerebral cortex can become specialized for different functions.
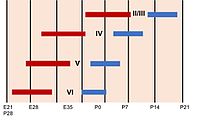
[58] During the period of cortical neurogenesis and layer formation, many higher mammals begin the process of gyrification, which generates the characteristic folds of the cerebral cortex.
[6] In contrast to the highly conserved circuitry of the medulla oblongata, for example, which serves critical functions such as regulation of heart and respiration rates, many areas of the cerebral cortex are not strictly necessary for survival.
[64] This theory states that new cortical areas are formed by the addition of new radial units, which is accomplished at the stem cell level.
The basal ganglia receive input from the substantia nigra of the midbrain and motor areas of the cerebral cortex, and send signals back to both of these locations.
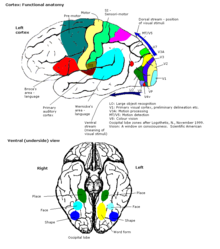
They function to produce a meaningful perceptual experience of the world, enable us to interact effectively, and support abstract thinking and language.
However, the processes of language expression and reception have been shown to occur in areas other than just those structures around the lateral sulcus, including the frontal lobe, basal ganglia, cerebellum, and pons.
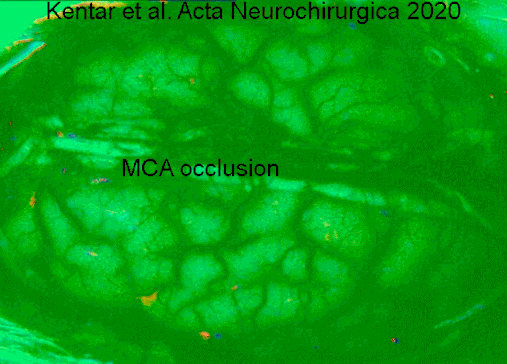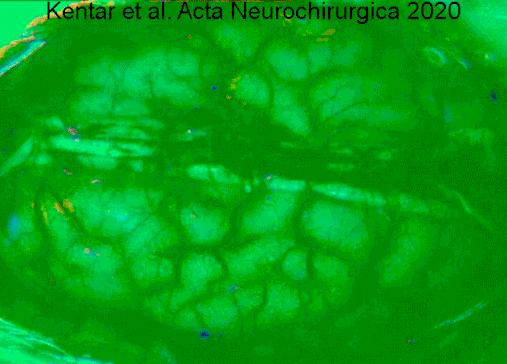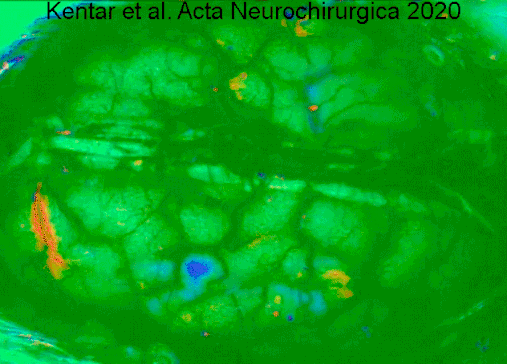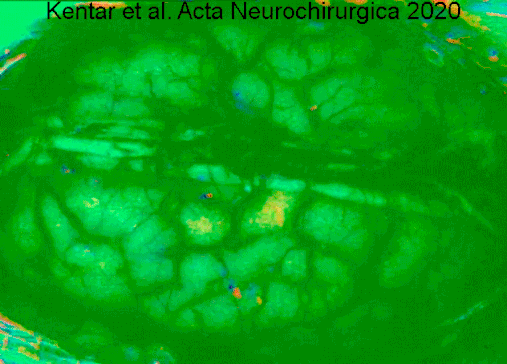
A type of electrocorticography called cortical stimulation mapping is an invasive procedure that involves placing electrodes directly onto the exposed brain in order to localise the functions of specific areas of the cortex.
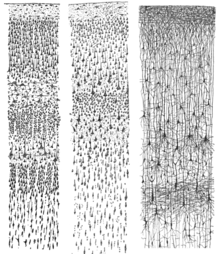
[82] Rafael Lorente de Nó, a student of Santiago Ramon y Cajal, identified more than 40 different types of cortical neurons based on the distribution of their dendrites and axons.
However, a study published in the journal Cell in 2010, based on gene expression profiles, reported strong affinities between the cerebral cortex and the mushroom bodies of the ragworm Platynereis dumerilii.