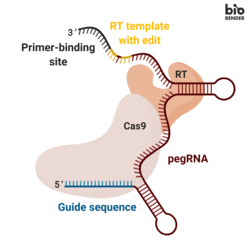
Prime editing
It mediates targeted insertions, deletions, and base-to-base conversions without the need for double strand breaks (DSBs) or donor DNA templates.
Once internalized, the fusion protein nicks the target DNA sequence, exposing a 3’-hydroxyl group that can be used to initiate (prime) the reverse transcription of the RT template portion of the pegRNA.
[1] In the first system, a wild-type Moloney Murine Leukemia Virus (M-MLV) reverse transcriptase was fused to the Cas9 H840A nickase C-terminus.
[1] In order to enhance DNA-RNA affinity, enzyme processivity, and thermostability, five amino acid substitutions were incorporated into the M-MLV reverse transcriptase.
To avoid this problem during DNA heteroduplex resolution, an additional single guide RNA (sgRNA) is introduced.
[9] Prime editor 4 utilizes the same machinery as PE2, but also includes a plasmid that encodes for dominant negative MMR protein MLH1.
Dominant negative MLH1 is able to essentially knock out endogenous MLH1 by inhibition, thereby reducing cellular MMR response and increasing prime editing efficiency.
[11][12][more detail needed] Prime editing was developed in the lab of David R. Liu at the Broad Institute and disclosed in Anzalone et al.
The prime editing technology has the potential to correct the vast majority of pathogenic alleles that cause genetic diseases, as it can repair insertions, deletions, and nucleotide substitutions.
This is an important feature of this technology given that DNA repair mechanisms such as NHEJ and HDR, generate unwanted, random insertions or deletions (INDELs).
Larger genetic alterations would require a longer RT template, which could hinder the efficient delivery of pegRNA to target cells.
[1][18] Prime editing in plants suffers from low efficiency ranging from zero to a few percent and needs significant improvement.
[9] Base editors used for prime editing require delivery of both a protein and RNA molecule into living cells.
Unfortunately, the effective packaging capacity of AAV vectors is small, approximately 4.4kb not including inverted terminal repeats.
[22] As a comparison, an SpCas9-reverse transcriptase fusion protein is 6.3kb,[1][23] which does not even account for the lengthened guide RNA necessary for targeting and priming the site of interest.
However, successful delivery in mice has been achieved by splitting the editor into two AAV vectors[2][3][4][24] or by using an adenovirus,[3] which has a larger packaging capacity.