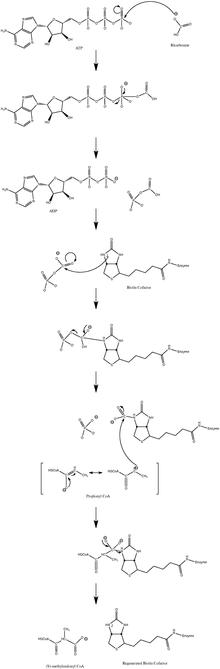
It is acted upon by a racemase, yielding (R)-methylmalonyl-CoA, which is then converted into succinyl-CoA by methylmalonyl-CoA mutase (one of the few metabolic enzymes which requires vitamin B12 as a cofactor).
[4] The BC and CT sites are approximately 55 Å apart, indicative of the entire BCCP domain translocating during catalysis of the carboxylation of propionyl-CoA.
[4] Site-directed mutagenesis at D422 shows a change in the substrate specificity of the propionyl-CoA binding site, thus indicating this residue's importance in PCC's catalytic activity.
[6] In 1979, inhibition by phenylglyoxal determined that a phosphate group from either propionyl-CoA or ATP reacts with an essential arginine residue in the active site during catalysis.
In future pregnancy studies, the use of lymphocyte PCC activity data should prove valuable in assessment of biotin status.
[15] When multiple copies of a polypeptide encoded by a gene form an aggregate, this protein structure is referred to as a multimer.
[16] This finding of intragenic complementation indicated that the multimeric structure of PCC allows cooperative interactions between the constituent PCCβ monomers that can generate a more functional form of the holoenzyme.
a. Carbamazepine (antiepileptic drug): significantly lowers enzyme levels in the liver[17] b. E. coli chaperonin proteins groES and groEL: essential for folding and assembly of human PCC heteromeric subunits[18] c. Bicarbonate: negative cooperativity[8] d. Mg2+ and MgATP2−: allosteric activation[19] a.
6-Deoxyerythronolide B: decrease in PCC levels lead to increased production [20] b. Glucokinase in pancreatic beta cells: precursor of beta-PCC shown to decrease KM and increase Vmax; activation [21] (See Template:Leucine metabolism in humans – this diagram does not include the pathway for β-leucine synthesis via leucine 2,3-aminomutase)
Figure 1.(a). Schematic drawing of the structure of the RpPCCα-RdPCCβ chimera, viewed down the three-fold symmetry axis. Domains in the α and β subunits in the top half of the structure are given different colors, and those in the first α and β subunits are labeled. The α and β subunits in the bottom half are colored in magenta and green, respectively. The red arrow indicates the viewing direction of panel b. (b). Structure of the RpPCCα-RdPCCβ chimera, viewed down the two-fold symmetry axis. The red rectangle indicates the region shown in detail in Fig. 2a. (c). Cryo-EM reconstruction of HsPCC at 15 Å resolution, viewed in the same orientation as panel a. The atomic model of the chimera was fit into the cryo-EM envelope. (d). The cryo-EM reconstruction viewed in the same orientation as panel b. The arrows indicate a change in the BCCP position that is needed to fit the cryo-EM map. All the structure figures were produced with PyMOL (www.pymol.org), and the cryo-EM figures were produced with Chimera.
[
5
]
This provides clear evidence of crucial dimeric interaction between alpha and beta subunits.
Figure 2.(a). Schematic drawing of the relative positioning of the BC and CT active sites in the holoenzyme. One α subunit and a β2 dimer (β1 from one layer and β4 from the other layer) are shown, and the viewing direction is the same as Fig. 1b. The two active sites are indicated with the stars, separated by 55 Å distance. The bound positions of ADP in complex with E. coli BC 18 and that of CoA in complex with the 12S subunit of transcarboxylase 21 are also shown. (b). Detailed interactions between BCCP-biotin and the C domain of a β subunit. Hydrogen-bonding interactions are indicated with the dashed lines in red. The N1′ atom of biotin is labeled as 1′, hydrogen-bonded to the main-chain carbonyl of Phe397. (c). Molecular surface of the CT active site, showing a deep canyon where both substrates are bound. (d). Schematic drawing of the CT active site.
[
5
]