Ribonuclease P
[1] Further, RNase P is one of two known multiple turnover ribozymes in nature (the other being the ribosome), the discovery of which earned Sidney Altman and Thomas Cech the Nobel Prize in Chemistry in 1989: in the 1970s, Altman discovered the existence of precursor tRNA with flanking sequences and was the first to characterize RNase P and its activity in processing of the 5' leader sequence of precursor tRNA.
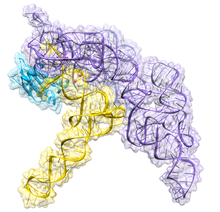
This crystal structure confirms earlier models of substrate recognition and catalysis, identifies the location of the active site, and shows how the protein component increases RNase P functionality.
Although the archaeal and eukaryotic holoenzymes have a much greater protein content than the eubacterial ones, the RNA cores from all the three lineages are homologous—helices corresponding to P1, P2, P3, P4, and P10/11 are common to all cellular RNase P RNAs.
As revealed by in vitro reconstitution experiments these protein subunits are individually dispensable for tRNA processing that is essentially mediated by the RNA component.
It has recently been argued that the archaebacterium Nanoarchaeum equitans does not possess RNase P. Computational and experimental studies failed to find evidence for its existence.
[23] RNase P is now being studied as a potential therapy for diseases such as herpes simplex virus,[24] cytomegalovirus,[24][25] influenza and other respiratory infections,[26] HIV-1[27] and cancer caused by fusion gene BCR-ABL.
[24][28] External guide sequences (EGSs) are formed with complementarity to viral or oncogenic mRNA and structures that mimic the T loop and acceptor stem of tRNA.