Succinate dehydrogenase
[2] In step 6 of the citric acid cycle, SQR catalyzes the oxidation of succinate to fumarate with the reduction of ubiquinone to ubiquinol.
The first two subunits, a flavoprotein (SDHA) and an iron-sulfur protein (SDHB), form a hydrophilic head where enzymatic activity of the complex takes place.
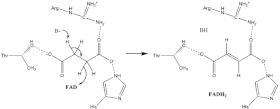
SDHA contains a covalently attached flavin adenine dinucleotide (FAD) cofactor and the succinate binding site and SDHB contains three iron-sulfur clusters: [2Fe-2S], [4Fe-4S], and [3Fe-4S].
[3] The subunits form a membrane-bound cytochrome b complex with six transmembrane helices containing one heme b group and a ubiquinone-binding site.
[4] Two distinctive ubiquinone binding sites can be recognized on mammalian SDH – matrix-proximal QP and matrix-distal QD.
This process is enhanced by succinate dehydrogenase assembly factor 2 (SDHAF2;[9] also called SDH5 in yeast and SDHE in bacteria) and by some of the Krebs cycle intermediates.
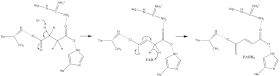
[10] Through studies of the bacterial system, the mechanism of FAD attachment has been shown to involve a quinone:methide intermediate.
[7] Fe-S prosthetic groups of the subunit SDHB are being preformed in the mitochondrial matrix by protein complex ISU.
Two more SDH assembly factors, SDHAF1 (SDH6) and SDHAF3 (SDH7 in yeast), seem to be involved in SDHB maturation in way of protecting the subunit or dimer SDHA-SDHB from Fe-S cluster damage caused by ROS.
A combination of mutagenesis and structural analysis identifies Arg-286 of the SDHA subunit (E. coli numbering) as the proton shuttle.
This facilitates a second hydrogen bond interaction between the O4 carbonyl group of ubiquinone and Ser27 of subunit C. Following the first single electron reduction step, a semiquinone radical species is formed.
His207 of subunit B is in direct proximity to the [3Fe-4S] cluster, the bound ubiquinone, and the heme; and could modulate electron flow between these redox centers.
The histidine residue decreases the pKa of tyrosine, making it more suitable to donate its proton to the reduced ubiquinone intermediate.
Succinate-analogue inhibitors include the synthetic compound malonate as well as the TCA cycle intermediates, malate and oxaloacetate.
In the 1980s simple benzanilides were found to have comparable activity to carboxin and a number of these were marketed, including benodanil, flutolanil and mepronil.
[14] More recently, other compounds with a broader spectrum against a range of plant pathogens have been developed including boscalid, fluopyram, fluxapyroxad, pydiflumetofen and sedaxane.
[19] The fundamental role of succinate-coenzyme Q reductase in both oxidative phosphorylation and the citric acid cycle makes it vital in all eukaryotic organisms.
[20] The penetrance data for SDH mutations causing tumorigenesis is lacking, and international guidelines suggest thorough screening for any carriers.
Buildup of succinate due to defective SDH can cause a pseudo-hypoxia and angiogenesis, both of which contribute to the distinctly vascular and characteristic "salt and pepper" appearance of paraganglioma on imaging.
This disruption in oxidative phosphorylation can lead to Leigh syndrome, mitochondrial encephalopathy, optic atrophy, myopathy, and a spectrum of disease.