Siglec
[2] Parallel studies by Ajit Varki and colleagues on the previously cloned CD22 (a B cell surface protein involved in adhesion and activation) showed direct evidence for sialic acid recognition.
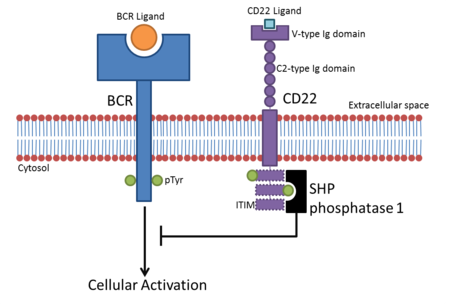
[7] These act to down-regulate signaling pathways involving phosphorylation, such as those induced by ITAMs (Immunoreceptor tyrosine-based activation motifs).
[6] This is best seen in Sialoadhesin, where arginine at position 97 forms salt bridges with the COO− group of the sialic acid, producing a stable interaction.
Sialic acid contains numerous hydroxyl groups which can be involved in the formation of glycosidic linkages, which are observed at carbons number 2, 3, 6, and 8 of the sugar backbone.
One exception is Sialoadhesin which contains 16 C2-Ig domains, producing a long, extended protein allowing it to bind trans-ligands, i.e. ligands found on other cells.
[10][11] Due to their ITIM-containing cytoplasmic regions, most Siglecs interfere with cellular signalling, inhibiting immune cell activation.
[12] The tyrosine contained within the ITIM is phosphorylated after ligand binding and acts as a docking site for SH2 domain-containing proteins like SHP phosphatases.
Siglec-14 appears to co-localise with Siglec-5, and as this protein inhibits cellular signalling pathways, co-ordinate opposing functions within immune cells.
Sialoadhesin lacks a cytosolic ITIM or a positive residue to bind ITAM-containing adaptors and so is thought not to influence signalling.
Studies show that this protein is involved in phagocytosis of bacteria that contain highly sialylated glycan structures such as the lipopolysaccharide of Neisseria meningitidis.
[29][30] The sialic acid family branches from glycans, sugar chains comprising various monosaccharides that cover the membrane of every living cell and display a staggering structural diversity.
As sialic acids are abundantly expressed in vertebrates and not in microorganisms, they are considered self-antigens or self-structures that play major role in inhibiting harmful immune system activity by regulating neutrophils and B cell tolerance.
[32][33] This evolutionary chain and incessant mutations have made Siglecs one of the most rapidly evolving gene, evidenced by both intra- and inter-species differences.
[34] The first attempts were made to develop high-affinity sialic acid mimetics for Siglec-2, which led to the discovery that with increased binding affinity came hydrogen bonding and lipophilic interactions between SAMs and Siglec-2.
[37] Success in drastically enhanced binding of SAMs to Siglec 2 suggests that a similar approach can work on other members of the family.
[39] Moreover, human cells, engineered with sialic acids carrying Ac5NeuNPoc incorporated into its sialoglycans and 3-bromo-benzyl azide, showed hyperactivity towards Siglec-2.