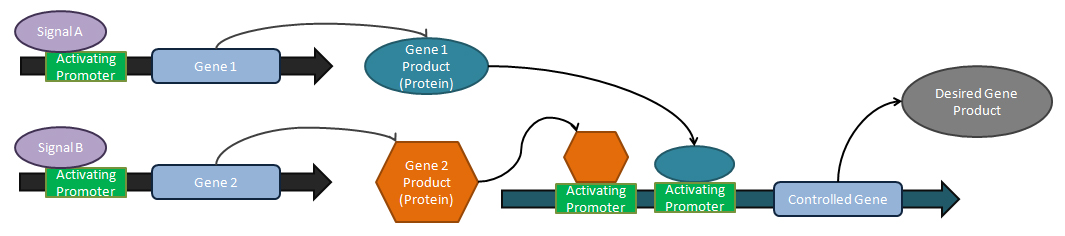
Synthetic biological circuit
In studies of diauxic growth of E. coli on two-sugar media, Jacques Monod and Francois Jacob discovered that E.coli preferentially consumes the more easily processed glucose before switching to lactose metabolism.
IPTG, a molecule similar to lactose, but with a sulfur bond that is not hydrolyzable so that E. coli does not digest it, is used to activate or "induce" the production of the new protein.
One, by Tim Gardner, Charles Cantor, and Jim Collins working at Boston University, demonstrated a "bistable" switch in E. coli.
[3] The second, by Michael Elowitz and Stanislas Leibler, showed that three repressor genes could be connected to form a negative feedback loop termed the Repressilator that produces self-sustaining oscillations of protein levels in E.
[5] There has been significant interest in encouraging education and outreach as well: the International Genetically Engineered Machines Competition[6] manages the creation and standardization of BioBrick parts as a means to allow undergraduate and high school students to design their own synthetic biological circuits.
[8] However, methods involving direct genetic introduction are not inherently effective without invoking the basic principles of synthetic cellular circuits.
This is a biological circuit where a simple repressor or promoter is introduced to facilitate creation of the product, or inhibition of a competing pathway.
However, with the limited understanding of cellular networks and natural circuitry, implementation of more robust schemes with more precise control and feedback is hindered.
Development in understanding cellular circuitry can lead to exciting new modifications, such as cells which can respond to environmental stimuli.
For example, cells could be developed that signal toxic surroundings and react by activating pathways used to degrade the perceived toxin.
[21] At the moment, circuit design is improving at a slow pace because of insufficient organization of known multiple gene interactions and mathematical models.

1 : RNA polymerase, 2 : Repressor, 3 : Promoter, 4 : Operator, 5 : Lactose, 6 : lacZ , 7 : lacY , 8 : lacA .