Type I topoisomerase
DNA topoisomerases regulate the number of topological links between two DNA strands (i.e. change the number of superhelical turns) by catalysing transient single- or double-strand breaks, crossing the strands through one another, then resealing the breaks.
There are several variants of Type IA topoisomerases, differing by appendages attached to the main core (sometimes referred to as the "topo-fold").
The enzyme uses the hydrolysis of ATP to introduce positive supercoils and overwinds DNA, a feature attractive in hyperthermophiles, in which reverse gyrase is known to exist.
Rodriguez and Stock have done further work to identify a "latch" that is involved in communicating the hydrolysis of ATP to the introduction of positive supercoils.
The topo III variant is likewise very interesting because it has zinc-binding motifs that is thought to bind single-stranded DNA.
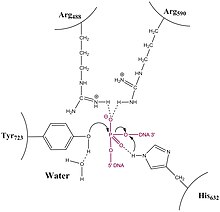
Relaxation is not an active process and energy (in the form of ATP) is not spent during the nicking or ligation steps; this is because the reaction between the tyrosine residue at the active site of the enzyme with the phosphodiester DNA backbone simply replaces one phosphomonoester bond with another.
No data suggest that Topo IB "controls" the swiveling insofar as that it has a mechanism in place that triggers religation after a specific number of supercoils removed.
On the contrary, single-molecule experiments suggest that religation is a random process and has some probability of occurring each time the swiveling 5'-OH end comes in close proximity with the attachment site of the enzyme-linked 3'-end.
All topoisomerases form a phosphotyrosine intermediate between the catalytic tyrosine of the enzyme and the scissile phosphoryl of the DNA backbone.
The active irinotecan metabolite, SN-38, acts by trapping (making a ternary complex with) a subset of Top1cc, those with a guanine +1 in the DNA sequence.
[11] One irinotecan-derived SN-38 molecule stacks against the base pairs flanking the topoisomerase-induced cleavage site and poisons (inactivates) the topoisomerase 1 enzyme.
This inhibition appears to be an adaptation to subtly modulate host topoisomerase I activity during infection to ensure optimal phage yield.
[14] In pre-clinical studies related to cancer, the deficient DNA repair genes include ATM[15] and NDRG1.
[16][17] Autoantibodies targeted against type I topoisomerase are called anti-Scl-70 antibodies, named by the association with scleroderma and the 70 kD extractable immunoreactive fragment that can be obtained from the otherwise larger (100-105 kD) target topoisomerase antigen (called the SCL-70 Antigen) of the antibodies.