Type VI secretion system
[8][9] While most of the early studies of Type VI secretion focused on its role in the pathogenesis of higher organisms, it is now known to function primarily in interbacterial antagonism.
[3] Studies have also shown that T6SS plays a role in the acquisition of essential metals, such as manganese and iron, from the surrounding environment.
This ability allows bacteria to outcompete rivals for these nutrients while fostering cooperation with related bacterial cells.
This suggests that T6SS plays a critical role in maintaining microbial community stability by balancing cooperation and competition.
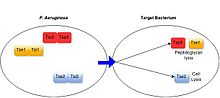
Members of the second class of substrates are targeted for secretion via interaction with the phage tail spike-like protein VgrG.
These substrates are often modular proteins, such as the Rhs toxins, that possess PAAR domain for interaction with VgrG at one end.
For example, the intracellular pathogen Francisella tularensis requires the activity of a T6SS to escape from phagosomes and replicate in the cytoplasm of macrophages.
[19] Burkholderia pseudomallei and Edwardsiella tarda are two other organisms which possess a T6SS that appears dedicated for eukaryotic targeting.
[20] A wide range of Gram-negative bacteria have been shown to have antibacterial T6SSs, including opportunistic pathogens such as Pseudomonas aeruginosa,[21] obligate commensal species that inhabit the human gut (Bacteroides spp.
[3][24] T6SS-positive bacterial species prevent T6SS-mediated intoxication towards self and kin cells by producing immunity proteins specific to each secreted toxin.
This system regulates the expression of Rsm small regulatory RNA molecules, and has also been implicated in biofilm formation.
LuxQ activates LuxU, which then acts on LuxO, a DNA-binding protein which represses HapR gene expression.