VLDL receptor
[5] This receptor has an important role in cholesterol uptake, metabolism of apolipoprotein E-containing triacylglycerol-rich lipoproteins, and neuronal migration in the developing brain.
Mutations of this gene may lead to a variety of symptoms and diseases, which include type I lissencephaly, cerebellar hypoplasia, and atherosclerosis.
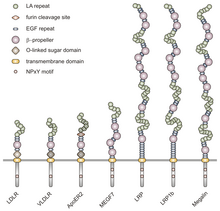
The NPxY motif functions in signal transduction and the targeting of receptors to coated pits and consists of the sequence Asparagine-Proline-X-Tyrosine, where X can be any amino acid.
[6] Mimicking this general structure, VLDLR has eight, 40 amino acid long cysteine-rich repeats in its extracellular N-terminal ligand-binding domain.
[9] The VLDLR O-linked glycosylation domain, next in the sequence, has many threonine and serine residues and totals 46 amino acids.
[7] Together, the exons making up the VLDLR gene encode a protein that is 873 amino acid residues long.
It has been shown that 75% of VLDLR transcripts exist as isoform type II in mouse brain models.
This shows that most VLDLRs in the brain are not glycosylated, as type II lacks exon 16 which encodes the O-glycosylation domain.
This binding of THBS1, however, does not induce the subsequent degradation of these transcription factors, as reelin does, and can thus lead to greatly amplified effects.
[7] In addition, it has been uncovered that this receptor is found, sub-cellularly, in the non-lipid raft sections of cell membranes.
Normal SRE-1 sequences, like those found in LDLR, are characterized by two repeats of the codon CAC separated by two intervening C nucleotides (5’-CACCCCAC-3’).
A 2010 study showed that the prescription drug Pioglitazone, an agonist of PPAR-γ, increases VLDLR mRNA expression and protein levels in experiments using mouse fibroblasts.
The Pioglitazone treated mice exhibited a higher conversion rate of plasma triglycerides into epididymal fats.
Thyroid hormone positively regulates VLDLR expression in skeletal muscles of rats, but not in adipose or heart tissues.
In rabbits, VLDLR expression in heart muscle is up-regulated by estrogen and down-regulated by granulocyte-macrophage colony-stimulating factor.
Despite this knowledge on its role in lipoprotein uptake, the complete mechanism of lipid metabolism performed by VLDLR is not fully understood.
At this point, hydrolysis occurs and lipoprotein is released into the cytoplasm while the receptors are recycled back to the cell surface.
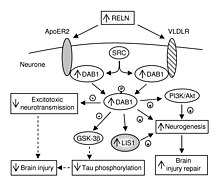
Finally, phosphorylated Dab1 activates an intracellular signaling cascade that directs neuroblasts to their proper location through the alteration of the cytoskeleton.
In addition to the organization of the neocortex, VLDLR also plays a role in neuronal migration of the hippocampus and the Purkinje cells of the cerebellum.
These disorders are usually associated with cholesterol homeostasis or a disorganization of neuron ordering in the brain due to disruption of the reelin pathway.
[7] In addition, being that apoE, a major ligand of VLDLR, is a leading genetic risk factor for Alzheimer’s disease, VLDLR may play a role in modulating the risk of this disorder [6] which is explained by the fact that a decrease in reelin signaling in the fascia dentata is supposed to initiate Alzheimer's disease.
[7] Type I lissencephaly, or agyria-pachygyria, is a rare developmental disorder characterized by the absence of gyri and sulci in the brain.
A homozygous deletion affecting the VLDLR gene results in a low degree of cortical thickening and absence of a cell-sparse zone.
[19] In a 2005 study, DES was renamed as VLDLR-associated cerebellar hypoplasia (VLDLRCH) after its cause was linked to a disruption in the VLDLR gene.
[20] At least six mutations affecting the homozygous recessive allele of the VLDLR gene have been identified and found to cause VLDLRCH.
One such mutation is a cytosine to thymine transition at base pair 1342 in exon 10 that causes a substitution at Arg448 for a termination signal.
Likewise, there is evidence of a cytosine to thymine transition at base pair number 769 in exon 5 that causes a substitution at Arg257 for a termination signal.
The recognized symptoms of VLDLRCH are moderate-to-severe intellectual disability, seizures, dysarthria, strabismus and delayed locomotion.
[22] Atherosclerosis is marked by an excessive accumulation of cholesterol by macrophages, leading to their transformation into foam cells.
Since macrophages do not have the ability to limit the influx of cholesterol, the balance is completely dependent on efflux pathways.