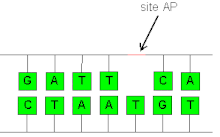
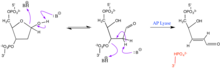AP site
A variety of glycosylases that recognize different types of damage exist, including oxidized or methylated bases, or uracil in DNA.
In alternative fashion, bifunctional glycosylase-lyases can cleave the AP site, leaving a 5' phosphate adjacent to a 3' α,β-unsaturated aldehyde.
The situation is more complex in higher eukaryotes, with different nucleotides showing a preference depending on the organism and experimental conditions.
Alkylation, deamination, and oxidation of individual bases can all lead to the weakening of the glycosyl bond, so exposure to agents that cause those modifications can encourage AP site formation.
Hydroxyl radicals can attack the glycosidic linkages, directly creating an AP site, or make the glycosyl bond less favorable by linking to the base or the deoxyribose ring.
[2] Enzymes, namely DNA glycosylases, also commonly create AP sites, as part of the base excision repair pathway.
Apyrimidinic sites form at a rate roughly 20 times slower, with estimates at around 500 formation events per day, per cell.
The single-stranded breaks occurring due to β-elimination require repair by DNA Ligase in order to avoid mutation.
[5] In calorie restricted rats, by comparison, AP endonuclease activity in these brain regions remains higher with age.
[5] These findings suggest that elevated AP site repair in calorie restricted animals may delay the aging process.