Meiotic recombination checkpoint
The main purpose of this step is to make sure that DNA replication has completed without error so that spindle pole bodies can separate.
The abrupt onset of M-Cdk in late prophase I depends on the positive transcription regulation feedback loop consisting of Ime2, Ndt80 and Cdk/cyclin complex.
[6][7] Under this cascade, failure to synapse will maintain the gene expression from sex chromosomes and some products may inhibit cell cycle progression.
Meiotic sex chromosome inactivation only happens in male, which may partially be the reason why only Spo11 mutant spermatocytes but not oocytes fail to transition from prophase I to metaphase I.
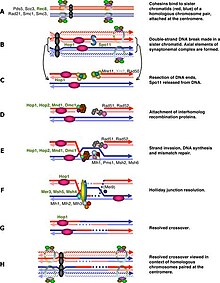
[11] During meiotic recombination in human and mouse, CHEK1 protein kinase is important for integrating DNA damage repair with cell cycle arrest.
[13] CHEK2 regulates cell cycle progression and spindle assembly during mouse oocyte maturation and early embryo development.
The response of oocytes to DNA double-strand break damage involves a pathway hierarchy in which ATR kinase signals to CHEK2 which then activates p53 and p63 proteins.
[citation needed] When a mutation inactivates Ndt80 in budding yeast, meiotic cells display a prolonged delay in late pachytene, the third stage of prophase.
[22][23] NDt80 is crucial for the completion of prophase and entry into meiosis 1, as it stimulates the expression of a large number of middle meiotic genes.
This protein is activated in late pachytene and is needed for crossover formation and partial loss of cohesion from chromosome arms.
[31] Heterokaryon Incompatibility (HI) has been likened to a fungal immune system;[32] it is a non-self recognition mechanism that is ubiquitous among filamentous members of the Asomycota phylum of the Fungi kingdom.
It has been found that mutations at the vib1 locus suppress non-self recognition, and VIB-1 is required for the production of downstream effectors associated with HI, such as extracellular proteases.
[36] Although usually characterized in yeast and other fungi, the DNA-binding domain of Ndt80 is homologous to a number of proteins in higher eukaryotes and the residues used for binding are highly conserved.
In humans, the Ndt80 homologue C11orf9 is highly expressed in invasive or metastatic tumor cells, suggesting potential usage as a target molecule in cancer treatment.