Cell junction
[6] Desmosomes, also termed as maculae adherentes, can be visualized as rivets through the plasma membrane of adjacent cells.
[7] Hemidesmosomes form rivet-like links between cytoskeleton and extracellular matrix components such as the basal laminae that underlie epithelia.
The band-type of adherens junctions is associated with bundles of actin filaments that also encircle the cell just below the plasma membrane.
Spot-like adherens junctions called focal adhesions help cells adhere to extracellular matrix.
[9] This is possible due to six connexin proteins interacting to form a cylinder with a pore in the centre called a connexon.
[9][10] Whilst variation in gap junction channels do occur, their structure remains relatively standard, and this interaction ensures efficient communication without the escape of molecules or ions to the extracellular fluid.
[11][12] Found in vertebrate epithelia, tight junctions act as barriers that regulate the movement of water and solutes between epithelial layers.
Tight junctions are classified as a paracellular barrier which is defined as not having directional discrimination; however, movement of the solute is largely dependent upon size and charge.
A three-dimensional image is still yet to be achieved and as such specific information about the function of tight junctions is yet to be determined.
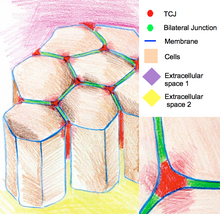
In some Drosophila epithelia, during cell divisions tricellular junctions establish physical contact with spindle apparatus through astral microtubules.
Tricellular junctions exert a pulling force on the spindle apparatus and serve as a geometrical clue to determine orientation of cell divisions.
[15] Selectins are cell adhesion molecules that play an important role in the initiation of inflammatory processes.
[19] Integrins act as adhesion receptors, transporting signals across the plasma membrane in multiple directions.
[20] Immunoglobulin superfamily are a group of calcium independent proteins capable of homophilic and heterophilic adhesion.