Cyclic nucleotide–gated ion channel
CNG channels have also been found to exist in prokaryotes, including many spirochaeta, though their precise role in bacterial physiology remains unknown.
In 1985, it was discovered that cGMP was able to directly activate the light-dependent response of rod ion channels by studying light-adapted retina of frogs.
[1] CNG channels were also found in cone photoreceptors, chemo sensitive cilia of olfactory sensory neurons, and the pineal gland.
After the identification of amino acids from purified proteins, cloning and functional expression of CNG channels were performed.
[4] CNG channels have important functions in signal transduction in retinal photoreceptors and olfactory receptor neurons.
Many studies have shown CNG channels in rod and cone photoreceptors, and they have also been found in the brain, the heart, the kidneys, and gonads.
[3] CNG channel homologs in Caenorhabditis elegans, Drosophila melanogaster, and Limulus polyphemus have unknown functions.
[5] [6] Spontaneous disulfide bond formation is state-dependent, implying that the conformational change in the helix bundle is affiliated with channel gating.
Small cations are able to move through an opening, which implies that the gate is beyond the helix bundle and that S6 helices are in conjunction with conformational changes in the selectivity filter.
Two CNG channel subunits, Tax-2 and Tax-4, have been cloned in C. elegans and are responsible for chemosensation, thermosensation, and normal axon outgrowth of some sensory neurons in C.
Changes induced by ligand binding occur in α helices (αA, αB, and αC and PBC helix).
A highly conserved residue of glutamic acid in the selectivity filter of CNG channels has been found to form a high-affinity binding site for Ca2+.
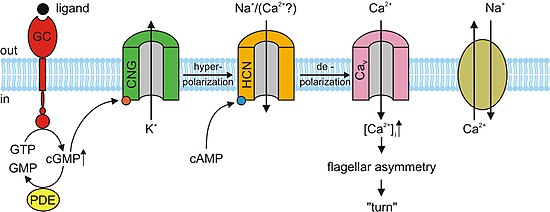
In the absence of cGMP, the CNG channels in the photoreceptors close preventing the flow of the aforementioned dark current.
This in turn causes a hyperpolarization of the outer segment of the photoreceptor, preventing the propagation of an action potential and the release of glutamate.
[11] Retinitis Pigmentosa (RP) is a genetic disease in which patients suffer degeneration of rod and cone photoreceptors.
The mutation of these subunits indirectly impairs rod cGMP-gated channel function, which implies that there is a common mechanism of photoreceptor degradation.
cAMP binds to the CNG channels in the OSN membrane, opening them, and making the cell highly permeable to Ca2+.
[3] cAMP and cGMP mediate several cellular responses such as acrosomal exocytosis, or the fusion of the male sperm to the female egg, and chemotaxis.
Although there has yet to be any establishment of a direct causal relationship, the previously mentioned observations suggest that cGMP activates calcium conductance.
Since A3 subunit knockout mice are fertile, CNG channels could be involved in some form of motility control and even in chemotactic swimming behavior or in the acrosomal exocytosis.
Male sterility can be accomplished by disrupting the CatSper1 gene; additionally, the cAMP-induced calcium influx is abolished in mutant mice.
It is possible that CNG and CatSper subunits assemble to form calcium-permeable and cyclic nucleotide-sensitive ion channels.
[3] cGMP-sensitive channels have been analyzed in the renal medulla, specifically in collecting duct cells, which influence the body's electrolyte and fluid balance.
[3] There has been identification of CNG ion channel subunits A2, A4, and B1 in a neuronal cell line that secretes gonadotropin-releasing hormone (GrH).
In high extracellular calcium, the unit conductance of CNG channels in rods and OSNs are significantly smaller than those measured in the neuronal line.
In plants, the Calmodulin binding domain (CaMBD) is found to overlap α-helix C in the CNBD of CNG channels.
[15] CNG channels play a large role in plant immunity and response to pathogens or external infectious agents.
CNG ion channels are also thought to be involved in pollen development in plants, however its exact role in this mechanism is still not known.
[16][17] Because these channels were only recently identified in spirochaeta and leptospira species,[16] their precise physiological function remains unknown in these organisms.
In order to develop these ideas, double electron-electron resonance (DEER) and rapid fixing techniques can show these mechanistic movements.