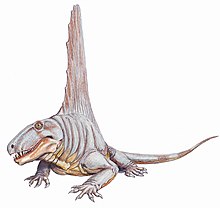
Dimetrodon
'two measures of teeth') is an extinct genus of sphenacodontid synapsid tetrapods that lived during the Cisuralian age of the Early Permian period, around 295–272 million years ago.
Most fossils have been found in the Southwestern United States, the majority of these coming from a geological deposit called the Red Beds of Texas and Oklahoma.
[9][10] Dimetrodon was a quadrupedal, sail-backed synapsid that most likely had a semi-sprawling posture between that of a mammal and a lizard and also could walk in a more upright stance with its body and the majority or all of its tail off the ground.
The second-largest species, D. grandis, has denticle serrations similar to those of sharks and theropod dinosaurs, making its teeth even more specialized for slicing through flesh.
[22] On the inner surface of the nasal section of the skull are ridges called nasoturbinals, which may have supported cartilage that increased the area of the olfactory epithelium, the layer of tissue that detects odors.
These ridges are much smaller than those of later synapsids from the Late Permian and Triassic, whose large nasoturbinals are taken as evidence for warm-bloodedness because they may have supported mucous membranes that warmed and moistened incoming air.
Since these first few caudal vertebrae narrow rapidly as they progress farther from the hip, many paleontologists in the late 19th and early 20th centuries thought that Dimetrodon had a very short tail.
The lower or proximal portion of the spine has a rough surface that would have served as an anchoring point for the epaxial muscles of the back and also has a network of connective tissues called Sharpey's fibers that indicate it was embedded within the body.
The cortical bone that grew over these breaks is highly vascularized, suggesting that soft tissue must have been present on the sail to supply the site with blood vessels.
[27] Layered lamellar bone makes up most of the neural spine's cross-sectional area, and contains lines of arrested growth that can be used to determine the age of each individual at death.
Their crookedness suggests that soft tissue may not have extended all the way to the tips of the spines, meaning that the sail's webbing may not have been as extensive as it is commonly imagined.
[35][36] The earliest discovery of Dimetrodon fossils were of a maxilla recovered in 1845 by a man named Donald McLeod, living in the British colony of Prince Edward Island.
Cope was the first to describe a sail-backed synapsid with the naming of C. natalis in his 1878 paper, although he called the sail a fin and compared it to the crests of the modern basilisk lizard (Basilicus).
In 1940, Romer coauthored a large study with Llewellyn Ivor Price called "Review of the Pelycosauria" in which the species of Dimetrodon named by Cope and Case were reassessed.
[63] Naosaurus would later be synonymized with Edaphosaurus, a genus which Cope named in 1882 on the basis of skulls that evidently belonged to herbivorous animals given their blunt crushing teeth.
[41] In 1940, Romer and Price recognized that the D. longiramus material belonged to the same taxon as another specimen described by paleontologist Samuel Wendell Williston in 1916, which included a similarly elongated mandible and a long maxilla.
[65] Romer and Price assigned Case and Williston's specimens to a newly erected genus and species, Secodontosaurus longiramus, that was closely related to Dimetrodon.
The modern view of synapsid relationships was proposed by paleontologist Robert R. Reisz in 1986, whose study included features mostly found in the skull rather than in the postcranial skeleton.
[27] In 1940, Alfred Romer and Llewellyn Ivor Price proposed that the sail served a thermoregulatory function, allowing individuals to warm their bodies with the Sun.
For example, in a 1973 article in the journal Nature, paleontologists C. D. Bramwell and P. B. Fellgett estimated that it took a 200 kilograms (440 lb) individual about one and a half hours for its body temperature to rise from 26 to 32 °C (79 to 90 °F).
[74] Most of these studies give two thermoregulatory roles for the sail of Dimetrodon: one as a means of warming quickly in the morning, and another as a way to cool down when body temperature becomes high.
Dimetrodon and all other Early Permian land vertebrates are assumed to have been cold-blooded or poikilothermic, relying on the sun to maintain a high body temperature.
Because of its large size, Dimetrodon had high thermal inertia, meaning that changes in body temperature occurred more slowly in it than in smaller-bodied animals.
Turner and Tracy noted that early therapsids, a more advanced group of synapsids closely related to mammals, had long limbs which can release heat in a manner similar to that of the sail of Dimetrodon.
The researchers concluded that the sail of Dimetrodon grew at a much faster rate than was necessary for thermoregulation, and suggested that sexual selection was the primary reason for its evolution.
[44] Fossils of Dimetrodon are known from the United States (Texas, Oklahoma, New Mexico, Arizona, Utah and Ohio), Canada (Prince Edward Island) and Germany, areas that were part of the supercontinent Euramerica during the Early Permian.
In addition to Dimetrodon, the most common tetrapods in the Red Beds and throughout Early Permian deposits in the southwestern United States, are the amphibians Archeria, Diplocaulus, Eryops, and Trimerorhachis, the reptiliomorph Seymouria, the reptile Captorhinus, and the synapsids Ophiacodon and Edaphosaurus.
These tetrapods made up a group of animals that paleontologist Everett C. Olson called the "Permo-Carboniferous chronofauna", a fauna that dominated the continental Euramerican ecosystem for several million years.
Dimetrodon was probably the top predator of the Red Beds ecosystem, feeding on a variety of organisms such as the shark Xenacanthus[citation needed], the aquatic amphibians Trimerorhachis and Diplocaulus, and the terrestrial tetrapods Seymouria and Trematops.
Insects are known from the Early Permian Red Beds and were probably involved to some degree in the same food web as Dimetrodon, feeding small reptiles like Captorhinus.