Hemozoin
Several currently used antimalarial drugs, such as chloroquine and mefloquine, are thought to kill malaria parasites by inhibiting haemozoin biocrystallization.
[22] However, studies on Schistosoma mansoni have revealed that this parasitic worm produces large amounts of hemozoin during its growth in the human bloodstream.
Although the shapes of the crystals are different from those produced by malaria parasites,[23] chemical analysis of the pigment showed that it is made of hemozoin.
[24][25] In a similar manner, the crystals formed in the gut of the kissing bug Rhodnius prolixus during digestion of the blood meal also have a unique shape, but are composed of hemozoin.
[26] Hz formation in R. prolixus midgut occurs at physiologically relevant physico-chemical conditions and lipids play an important role in heme biocrystallization.
Heme is retained in the peritrophic matrix, a layer of protein and polysaccharides that covers the midgut and separates gut cells from the blood bolus.
[28] Although β-hematin can be produced in assays spontaneously at low pH, the development of a simple and reliable method to measure the production of hemozoin has been difficult.
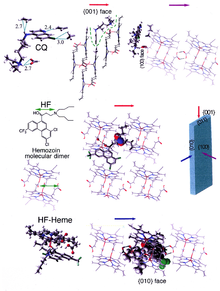
[30] β-Hematin crystals are made of dimers of hematin molecules that are, in turn, joined together by hydrogen bonds to form larger structures.
[33][34] They also exhibit optical dichroism, meaning they absorb light more strongly along their length than across their width, enabling the automated detection of malaria.
[36] Hemozoin formation is an excellent drug target, since it is essential to malaria parasite survival and absent from the human host.
[40][41] Hemozoin is released into the circulation during reinfection and phagocytosed in vivo and in vitro by host phagocytes and alters important functions in those cells.
[44][45][46] In contrast, a powerful, short-term stimulation of oxidative burst by human monocytes was also shown to occur during phagocytosis of nHZ.